- Entsyymin cPLA2 merkitys aivossa
Tekstissä käyttettyjä lyhennyksiä
Fosfolipaasit A2 ( EC 3.1.1.4; PLA2)
PLA2 voidaan luokitella 11 eri tyyppiin
Mitä PLA2 tekee?
Poistogeenitutkimus
Mitä tuloksia saatiin?
Esteröitymättömät rasvahapot (NEFA)
Aivojen fosfolipidit (PL)
Esteröityneet rasvahapot
Arakidonihappo , AA, C20:4 n6
cPLA2 puutteen merkitys: arakidonihapon menetystä
Muita lipolyyttisiä entsyymeitä ilmenee aivoissa
cPLA selektiivisempi arakidonihapolle (AA) kuin dokosahexaanihapolle (DHA)
vakutus aivojen funktioon ja struktuuriin?
Miksi poistogeeniset hiiret olivat resistenttejä aivoarteriatukokselle?
Erot aivojen esteröityneitten PUFA happojen pitoisuuksissa
Lajiero
Entsyymin cPLA2 merkitys aivossa
http://www.jlr.org/cgi/content/full/44/1/109
2003 by Lipid Research, Inc.
Thad A. Rosenberger et al. Brain lipid metabolism in the cPLA2 knockout mouse Journal of Lipid Research, Vol. 44, 109-117, January 2003 Brain Physiology and Metabolism Section, National Institute on Aging, National Institutes of Health, Bethesda, MD 20892
Harvard Medical School, Charlestown.
Tässä on tutkimuksen kohteena aivojen fosfolipidien (PL) metabolia hiiressä.
Normaali sytosolinen fosfolipaasi P2 (cPLA2) oli poistogeenisena tutkittavana myös. Kun verrattiin normaaligeenistä ja poistogeenistä havaitiin muutoksia useissa aivojen fosfolipidipitoisuuksissa, esteröitynyttä linolihappoa (C18:2 n6) , arakidonihappoa (C20:4 n6) ja dokosahexaeenihappoa( C22:6 n3) oli muuntuneet määrät koliiniglyserofosfolipideissä (PC). Vähentyneitä määriä esteröitynyttä arakidonihappoa (C20:4) oli fosfatidyylinositolissa(PI).
http://www.aapsj.org/articles/aapsj0802/aapsj080236/aapsj080236_figure2.jpg
{kind=link}
Poistogeenisen hiiren arakidonihappojen (AA) siirtyminen plasmasta ja aivojen arakidonyyliCoA aitiosta etanolaminiglyserofosfolipidiin (EtnGpl) ja koliiniglyserofosflipidiin (ChoGpl) oli alentunut, mutta siirtymä fosfatidyylinositoliin (PI) oli kohonnut.
Nämä erot olivat korrelaatiossa aivon arakidonihapon muuntuneeseen turn over prosessiin ja metabolisiin menetyksiin. Nämä tutkimukset osoittivat että sytosolinen entsyymi cPLA2 on välttämätön ylläpitämässä aivojen fosfolipidien ja niihin esteröityneitten PUFA rasvahappojen pitoisuuksia.
Huom: Alentuneet esteröityneen arakidonihapon (AA) ja dokosahexaeeniihapon (DHA) määrät saattavat selittää, miksi poistogeenisella hiirellä oli vastustuskykyä Arteria- media aivovaltimon tulpan muodostusta kohtaan ; seikka saattaa vaikuttaa kalvojen fluiditeettiin, neuroinflammaatioon, signaalien johtumiseen ja muihin aivoprosesseihin.
Tekstissä käytettyjä lyhennyksiä
AA, arachidonic acid; C20:4 n3
CerP Cho, sphingomyelin
ChoGpl, choline glycerophospholipid;
DHA, docosahexaenoic acid; C22:6 n3,
DG, diacylglycerol;
EtnGpl, ethanolamine glycerophospholipid;
PAF, platelet activating factor;
PLA2, phospholipases A2;
cPLA2, cytosolic phospholipase A2;
PlsCho, plasmenylcholine;
PtdCho, phosphatidylcholine;
PlsEtn, plasmenylethanolamine;
PtdEtn, phosphatidylethanolamine, PE, kefaliini
PtdIns, phosphatidylinositol; PI, lipositoli
PtdSer, phosphatidylserine, PS
PUFA, polyunsaturated fatty acid;
sPLA2, secretory phospholipase A2;
TG, triacylglycerol
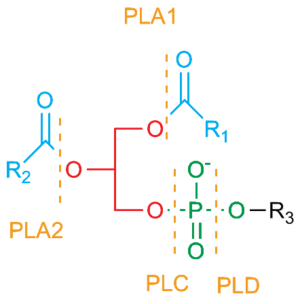
Fosfolipaasit A2 ( EC 3.1.1.4; PLA2)
Nämä entsyymit hydrolysoivat ( hajoittavat) rasvahappoja fosfolipidien (PL) sn2 asemasta ja muodostavat esteröitymättömiä rasvahappoja ( NEFA) ja lysofosfolipidejä (LPL).
http://upload.wikimedia.org/wikipedia/commons/thumb/e/e2/Phospholipases2.png/300px-Phospholipases2.png
{kind=link}
PLA2 voidaan luokitella 11 eri tyyppiin
perustaen niitten katalyyttiseen aktiivisuuteen, aminohappohomologiaan ja mRNA pilkkoutumisvariantteihin.Tyypit I, II, III, V, IX, X ja XI ovat matalamolekyylipainoisia ja ne vaativat kalsiumia Ca++ aktiivisuuteensa ja niillä on histidiini katalyyttisessä kohdassa. Tyypit IV, VI, VII, ja VIII ovat korkea molekyylipainoisia ja ne sijaitsevat solun sytosolissa. Nämä entsyymit käyttävät seriiniä katalyyttiseen aktiivisuuteensa ja kalsiumin riippuvuutta voi olla tai ei ole.
Nisäkäsaivoissa mRNA pitoisuudet osoittavat, että PLA2 tyypit IIA, IIC, IV ja VI PLA2 esiintyvät laajalti, kun taas tyyppi V PLA2 lmenee matalapitoisena muualla paitsi hippocampuksessa. Ihmisaivojen kyky modifioida eri PLA2 isoformien ilmenemistä antaa tiettyä riittävää kattavuutta rasvahappojen ja fosfolipidien metabolian säätelyyn.
Mitä PLA2 tekee?
PLA2 aktivaatiosta voi kalvon fosfolipidistä vapautua arakidonihappoa (AA; 20:4 n-6) tai dokosahexzaanihappoa(DHA; 22:6 n-3) sn-2 asemasta aivojen GLIA-SOLUISTA tai NEURONEISTA. Aktivaatio voi tuottaa myös lyso-PAF ( lyso-platelet activating factor), joka muuttuu PAF- muodoksi, kun sn2 asemaan esteröityminen tapahtuu. PAF on voimakas neurotransmitteriaine.
PLA2 voi myös aktivoitua kalvon G-proteiinilla, kun tiettyjä neurotransmitteriaineita kiinnittyy tiettyihin neuroreseptoreihin tai kun aktiivi kalsium Ca++ menee sisään soluun glutamiinihapon(glu) kiinnityttyä NMDA-reseptoriin
http://www.bris.ac.uk/Depts/Synaptic/info/glutamate.html
tai asetylkoliinin(ak) kiinnityttyä nikotiinireseptoriin.
Koska aivoissa eri PLA2-isoformit eivät toimi siten, että ne voisivat korvata toisiaan, poistogeenistrategioilla voidaan identifioida niiden erillisiä tehtäviä.
Poistogeenitutkimus
Tässä työssä tutkijat selvittivät aivojen lipidimetaboliaa poistogeenisellä hiirellä ( poissaoleva geeni vastasi 85 kDa tyyppi IV A sytosolista PLA2, entsyymiä) Tämä cPLA2 on selektiivinen arakidonihapolle(AA) enemmän kuin muille rasvahapoille ja vaatii sekä aktiivi kalsiumia(Ca++) että fosforylaation täyteen aktiivisuuteensa. Se on osallisena neuroreseptorin signaalin aloittamisessa ja sen transkriptiota säätää alas rotan aivossa LiCl, litiumkloridi.
Tällaisen (cPLA2) poistogeenisen hiiren neurologinen kehitys oli normaali. Naaraan reproduktiivinen kyky oli alentunut, kun taas koiraat olivat kontrolleita resistentimpiä arteria cerebri median tukosta vastaan. Poistogeeninen oli myös resitentimpi toksista molekyyliä MPTP vastaan, mitä indusoitui dopamiinin puutteessa ja se toipui nopeammin allergeenin indusoimasta bronkokonstriktiosta. Poistogeenisen hiiren peritoneaaliset makrofagit ja mastsolut eivät pystyneet tuottamaan eikosanoideja vasteena stimulaatioon , mikä viittasi vajaaseen AA kaskadiin ( eikosanoidikaskadiin). Kun näin ollaan identifioitu muutoksia aivojen lipidien metaboliassa poistogeenisellä hiirellä, voidaan paremmin ymmärtää, mikä rooli cPLA2 entsyymillä voi olla aivojen struktuurissa ja funktiossa.
Mitä tuloksia saatiin?
- Esteröitymättömät rasvahapot (NEFA)
Plasman ja aivojen esteröitymättömistä rasvahapoista (NEFA) ja aivojen aktivoiduista rasvahapoista (acylCoA)
Ei havaittu mitään statistisesti merkitsevää eroa tutkitun poistogeenisen ja kontrollihiiren plasman ja aivojen esteröitymättömien rasvahappojen pitoisuuksissa . Ei ollut myöskään mitään merkitsevää eroa minkään aktivoidun pitkäketjuisen rasvahapon ( LCFACoA) keskimääräisen pitoisuuden välillä näissä kahdessa ryhmässä. Ei myöskään AA-CoA aktiivisuuden ja plasman AA spesifisen aktiivisuuden suhde eronnut merkitsevästi näissä eri koe-eläimissä. Noin 4% aktivoituneesta arakidonihaposta on peräisin plasmasta ja 96% on vapautunut fosfolipideistä.
- Aivojen fosfolipidit (PL)
kun taas keskimääräinen pitoisuus seuraavia: EtnGpl ja ChoGpl eivät eronneet kontrolliarvoista. Keskimääräiset pitoisuudet seuraavia: PtdEtn, PlsEtn, PtdCho, PlsCho fraktiot eivät myöskään eronneet ryhmien kesken.
- Esteröityneet rasvahapot
Lipositoli (PI, PtdIns):
esteröitynyt AA oli alentunut 368 nmol/g (35%), kun taas esteröitynyt steariinihappo (C18:0) oli lisääntynyt 459 nmol/g. ja esteröitynyt palmitiinihappo (C16:0 ) oli lisääntynyt 70 nmol/g. ilmeisesti kompensoiden arakidonihapon (C20:4- n6) vähenemistä.
Fosfatidyyliseriini (PS,PtdSer)
osoitti vain yhtä merkitsevää eroa: steariinihappo(C18:0) oli lisääntynyt 30%.
ChoGpl,
koliiniglyserofosfolipideissä havaittiin eroa kaikissa esteröityneissä rasvahapoissa paitsi öljyhapossa ( C18:1). Esteröitynyt linolihappo(C18:2-n6) oli vähentynyt 99nmol/g (36%), arakidonihappo oli vähentynyt 671 nmol/g( 59%), DHA( C22:6 n39 oli vähentynyt 724 nmol/g (66%) . Koliiniglyserofosfolipideissä havaittavat erot poistogeenisten ja kontrollien välillä lienevät johtuneet lesitiinin eroista (PtdCho, PC) koska mitään merkitsevää eroa ei ollut plasmenyyli koliinissa PlsCho.
Arakidonihappo , AA, C20:4 n6
Arakidonihapon inkorporoituminen ja vaihtuminen yksittäisissä aivojen fosfolipideissä (PL)
Poistogeenisessa hiiressa havaittu merkitsevä esteröityneen arakidonihapon C20.4 n6 vähenemä aivojen inositolissa(PtdIns) johti plasmasta käsin otetun arakidonihapon inkorporaation nousuun. Tämä viittaa siihen, että AA tasapainon menetys aivojen inositolifosfolipideissä (PI) kiihtyy. Arakidonyyli CoA–aitiosta otetaan nyt lisääntynyt määrä AA rasvahappoa ja AA vaihtuminen ( turn over) lisääntyy kaksinkertaiseksi.
Vaikka arakidonihapon inkorporaatiotahti koliiniglyserofosfolipideihin (ChoGpl) plasmasta ja AA-CoA aitiosta aleni poistogeenisella hiirellä, arakidonihapon turn over koliiniglyserofosfolipideissä (ChoGpl) kuitenkin kohosi heijastaen epäsuhtaista arakidonihapon esteröitymistä. Kohonneita määriä havaittiin fosfatidyylikoliinissa (lesitiinissä, PC), mutta ei plasmenyylikoliinifraktiossa (PslCho) näissä koliiniglyserofosfolipideissä, joissa arakidonihapon vaihtuminen itse asiassa on alentunut.
Arakidonihapon inkorporaatiotahti etanolaminiglyserofosfolipideihin (EtnGpl) sekä plasmasta että aivon arakidonyyliCoA aitiosta oli alentunut 3 kertaisesti poistogeenisessa hiiressä , mikä pätee vahvasti alentuneeseen turnover prosessiin . Tämä vähenemä heijastui siten myös fosfatidyylietanolaminin eli kefaliinin (PtdEtn, PE) vaihtumiseen , koska PlsEtn plasmenyylietanolaminifraktion turn over oli lisääntynyt.
HUOLIMATTA huomattavista aivojen fosfolipidien(PL) ja rasvahappokokoomusten ja arakidonihapon kinetiikan eroista aivojen eri fosfolipideissä, niin poistogeeninen hiiri ei osoittanut mitään merkitsevää eroa kontrollin kanssa plasman tai aivojen non-esteröitymättmien(NEFA) rasvahappojen konsentraatiossa, aivojen aktivoituneitten rasvahappojen (acylCoA) aivopitoisuuksissa. ja aivojen aktivoidun arakidonihapon laimentumistekijässä (dilution factor).
Jos olisi tyydytty tutkimaan vain non-esteröityneitten rasvahappojen plasma-ja aivo-pitoisuudet tai aivojen aktivoidun arakidonihapon ( acylCoA) pitoisuus, ei olisi saatu näyttöä huomattavasta lipidikomposition muuntumisesta ja arakidonihapon kinetiikasta poistogeenisellä hiirellä. Ainoa näyttö plasman muuntuneesta arakidonihappometaboliasta oli nopeampitahtinen merkatun arakidonihapon katoama infusoitaessa tritiumilla merkattua arakidonihappoa (35% lyhyempi puoliintumisaika plasmassa).
Rasvahappojen kinetiikan tutkimuksissa nähtiin lisääntynyt esteröitymättömän arakidonihapon (AA) inkorporaatio plasmasta ja aivon aktiivista arakidonihappoaitiosta (AA-CoA) fosfoatidyyliinositoliin (PI) poistogeenisessä hiiressä ja samalla lisääntynyt arakidonihapon turnover lipositolissa (PI, fosfatidyyli-inositolissa) että koliiniglyserofosfolipideissä (ChoGpl) huolimatta siitä, että esteröityneen arakidonihapon pitoisuudet olivat alemmat.
cPLA2 puutteen merkitys: arakidonihapon menetystä
Selvästikin PLA2 entsyymin puuttuminen johti lisääntyneeseeen arakidonihapon metaboliseen menetykseen, joko konversiolla eikosanoideiksi tai muuta metabolista tietä.
cPLA osallistuu esteröityneen arakidonihapon ylläpitoon fosfatidyyli-inostoleissa (PI) ja koliiniglyserofosfolipideissä.
Kun poistogeenisessa hiiressä kuitenkin vielä tapahtuu lisääntyvää AA inkorporaatiota ja vaihtumista fosfatidyyli-inositolissa(PI) ja vaihtumista koliiniglykofosfolipoideissä, vaikka esteröityneen arakidonihapon määrä on vähentynyt, tämä viittaisi siihen, että muut PLA2-isoformit voinevat kompensoida puuttuvan cPLA2 entsyymin aktiivisuutta säätelemällä näiden fosfolipidiluokkien aineenvaihduntaa.
Todellakin cPLA2 kuten tyypit IIA tai V sPLA2 voivat vahvistaa välitöntä tai viivästynyttä arakidonihapon vapautumsita vasteena SYTOKIINEILLE tai vasikan fetaaliseerumille.
Erilaiset PLA2 isoformit voivat myös hydrolysoida arakidonihappoa fosfolipideistä eri fosfolipidiluokille selektiivisellä tavalla.
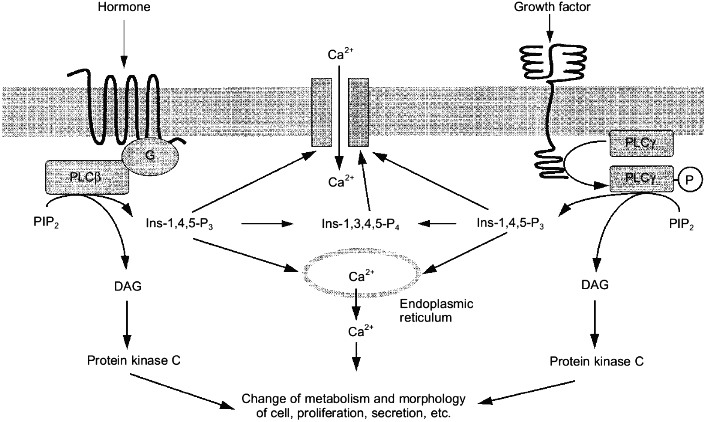
Muita lipolyyttisiä entsyymeitä ilmenee aivoissa
Fosfotidyyli-inositolille on spesifinen entsyymi PLC, fosfolipaasi C.
http://protein.bio.msu.ru/biokhimiya/contents/v63/full/63010047Fig2.gif
{kind=link}
Fosfatidyylikoliinille (PtdCho, PC=) on spesifinen entsyymi PLD, fosfolipaasi D.
Sen takia on entsymaattista riittävyyttä, koska on multippeleita PLA2 isoformeja ja/ tai kompensaatiota muista lipolyyttisistä entsyymeistä, joita aivoissa esiintyy, kun aivojen PtdIns ja ChoGpl turn over kohoaa poistogeenisessä hiiressä.
cPLA selektiivisempi arakidonihapolle (AA) kuin dokosahexaanihapolle (DHA)
Vaikka cPLA olisi selektiivisempi AA:lle kuin DHA:lle, sen jatkuva poissaolo vaikuttaa kuitenkin myös DHA hapon esteröitymisen huomattavaa vähenemistä eikä vain AA esteröitymisen vähenemistä. Näiden rasvahappojen metaboliset tiet ovat interaktiiviset ja ne säätyvät cPLA-entsyymillä osittain.
ESIMERKIKSI.
Se arakidonihapon eikosanoideiksi muuttuminen mikä tapahtuu 2-LO ja 5-LO ( lipoxygenaasi) syklo-oxygenaaseilla inhiboituu DHA –haposta.
On kilpailua aktivoitujen omega6 ( n6) ja omega3( n3) linjojen PUFA acylCoA happojen kesken entsyymeistä, jotka saturoivat( lisäävät kaksoissidoksia) ja elongoivat ( pidentävät) . Jos on dietääriä puutetta omega3- rasvahapoista, silloin aivojen arakidonihappo(AA) muuttuu n6 linjalla eikosapentaeenihapoksi (C22:5 n6).
Kuitenkin esteröitynyttä 22:5n-6 ei esiinny retrogradisen konversion puuttuessa
tai AA ketjun pitkittymä aivoissa poistogeenisellä ja kontrollihiirellä on samanlaisia kuin normaalipitoisuudet aivojen asyyliCoa ja plasman NEFA pitoisuuksissa.
Interaktio AA ja DHA aineenvaihdunnan välillä ei näytä tapahtuvan aivojen fosfolipidien välisenä kiertona. Tässä suhteessa jos koe-eläimiltä puuttuu n3- linjan PUFA kolmessa sukupolvessa, niillä on alentunut DHA turn over, mutta normaali AA turn over aivojen fosfolipideissä.Toisaalta taas krooninen litiumkloridin anto koe-eläimille vähensi AA vaihtuvuutta vaikuttamatta DHA vaihtuvuuteen.
Arakidonihapon ja Dokosahexaanihapon itsenäinen kierto fosfolipideissä on konsistenttia, kun on läsnä arakidonihapolle spesifinen PLA2 ja arakidonihapolle spesifinen aktivoidun arakidonihapon syntetaasi (Arachidonoyl CoA synthetase).
Poistogeenisessä hiiressä toisaalta muita fosfolipidejä metaboloivat entsyymit eri asyyliketjuspesifiteetein voivat vaikuttaa havaittuja muutoksia sekä AA että DHA kompositioissa. Lisätutkimuksia tarvitaan cPLA entsyymin puuttumisen merkityksestä niihin entsyymeihin ja niiden kyvystä pitää yllä plasman esteröitymättömät rasvahapot (NEFA) ja lambda-AA CoA vaikka arakidonihapon puoliintumisaika ja aivojen fosfolipidien vaihtumistahti muuttuisi. Lisäksi tulisi määrittää DHA:n inkorporoituminen ja vaihtumistahti aivojen fosfolipideissä poistogeenisellä hiirellä. Mikä osuus on poistogeenisyydellä ja mikä taas kompensoivilla entsyymeillä?
Vaikutus aivojen funktioon ja struktuuriin?
Luulisi, että huomattava alenema esteröityneen linolihapon (C18:2n-6), arakidonihapon ja DHA:n määrissä koliiniglyserolifosfolipideissä ja arakidonihapon vähenemä fosfatidyyli-inositolissa aivojen fosfolipidien rakenteessa voisi heijastua muuntuneeseen aivofunktioon ja struktuuriin.
Vaikka poistogeenisessa hiiressä onkin sanottu neurologisen kehityksen pysyvän normaalina, sofistiset muistin ja käytöksen testit ovat tekemättä näistä koe-eläimistä. Jos ei ole muutoksia aivojen funktiossa, havaittujen muutosten merkitys aivojen normaalille fysiologialle ei ole selvä ( cPLA entyymin osallistuminen ja relevanssi aivojen fosfolipidien metaboliassa)
Miksi poistogeeniset hiiret olivat resistenttejä aivoarteriatukokselle?
Syy voi olla vapautuneen ja esteröityneen arakidonihapon vähempi saatavuus insultin aikana johtuen cPLA aktiivisuuden puutteesta. Sama löydös pätee myös kroonisesti administroituun Litiumkloridiin- Tämä nosti vastustuskykyä aivoperäistä iskemiaa kohtaan ja alensi aivojen cPLA2 expressiota ja arakidonihapon turn overia (siis oli aivoprotektiivinen).
Erot aivojen esteröityneitten PUFA happojen pitoisuuksissa
Poistogeenisillä odotetusti olisi odotettavissa muutoksia aivojen solukalvojen fluiditeetissa, reseptorien toiminnassa, kalvojen remodelling prosessissa, neuroplastisuudessa ja apoptoosiresistenssissä muun muassa.
cPLA2 entsyymin puuttuminen voi vaikuttaa reseptorisignaalin aloitusprosessiin, jossa se osallistuu vapauttamaan arakidonihappoa(AA) ja aloittamaan sen kaskadin.
Poistogeeniseltä raportoitu stimuloitujen makrofagien ja mastsolujen eikosanoidien muodostumisen puuttuminen sopii tähän oletukseen. Poistogeenisellä naaraalla tavataan alentunut fertiliteetti ja siihen voi olla syynä muutokset arakidonihappokaskadissa. Koska fetoplasentaalinen kehitys vaatii cPLA2 aktivaatiota ja prostaglandiinien muodostumista. Ylläolevien seikkojen takana voi olla cPLA aktiviteetin puuttuminen.
Mutta mikä on primäärin poistogeenisyyden ja sekundäärin kompensatiivisen mekanismin osuus, se on vielä selvittämättä. Mitä jatkotutkimuksissa tulisi ottaa huomioon? ( käytösmuutokset, muitten aivorasvahappojen vaihtuminen, myös DHA:n vaihtuminen, PtdIns ja PtdCho aineenvaihdunnan entsyymien ilmenemistavat, jotta saadaan parempi käsitys cPLA.n roolista aivoissa.
Lajiero
Etanolaminiglykofosfolipideissä (EtnGpl) oli arakidonihapon vaihtuminen kymmenen kertaa suurempi kontrollihiirissä kuin kontrollirotissa mikä on ehkä lajiero. Kontrollihiirillä oli vajetta ryhmän IIA sPLA2 entsyymistä. sPLA2 entsyymin spesifinen substraatti saattaa olla fosfatidyletanolamini(PE). tai sitten ero heijasti hiiren myeliinin valkean aineksen pienempää fraktiota verrattuna rotan aivon valkeaan ainekseen ja EtnGpl:n nopeampaa vaihtumista harmaassa aineessa kuin valkoisessa aineessa.
YHTEENVETO
Poistogeeninen eläin ei anna selkeää ymmärtämystä spesifisen geenituotteen normaalista roolista, koska sen fenotyyppi voi johtua primääristä geenin puutteesta aivan yhtä hyvin kuin sekundäärisistä muutoksista kehityksen ja kypsymisen aikana. Tässä suhteessa 50% reduktio aivon cPLA2 transkriptiossa aikuisrotalla (kroon.LiCl administraatio) ei muuntanut esteröityneitä arakidonihappojen ja DHA happojen aivopitoisuuksia , kun taas alensi arakidonihapon ( mutta ei DHA:n) vaihtumista aivojen fosfolipideissä, mistä voisi päätellä, että cPLA knockoutia osin muistuttava tila ( conditional knockout) voisi paremmin selvittää cPLA2 roolia kuin kokoiän poistogeenisyys. Siitä huolimatta cPLA2 poissolo koko kehityksen ja kypsymisen aikana johti moniin muutoksiin aivojen fosfolipidien kompositiossa, aivojen esteröityneitten arakidonihappojen ja DHA happojen redusoituneisiin määriin ja kinetiikan muuntumisiin aivojen arakidonihappojen aineenvaihdunnassa.
Lopuksi nämä tulokset osoittavat monia tapoja joilla cPLA poistogeeninen fenotyyppi voidaan edelleen tutkia, jotta cPLA:n säätelevä rooli aivojen struktuurille ymmärrettäisiin paremmin, ja miten se moduloi signaalinvälitystä ja myös mikä on sen rooli vaurioon liittyvissä mekanismeissa.
Nevertheless, the absence of cPLA2 throughout development and maturation results in multiple changes involving brain membrane phospholipid composition, reductions in the levels of brain esterified AA and DHA, and alterations in the kinetics of brain AA metabolism. Finally, these results identify multiple ways in which the cPLA2-/- phenotype can be further examined so as to better understand how cPLA2 regulates brain structure, modulates signal transduction, as well as aid in future studies investigating its role in mechanisms associated with injury. , 2002.
Artikkeli on vuodelta 2002
{kind=link}
At any rate, I liked some of the vadlo mouse cartoons!
SvaraRadera