http://www.avoin.helsinki.fi/oppimateriaalit/ravitsemustieteen_perusteet/04_rasvat_rakenne.shtml
Tässä ravitsemustieteen perusteessa huomaa rasvahappojen joukossa kasvisperäisen linolihapon.
Siitä muodostuu kehossa arakidonihappoa, jota joka solu lataa fosfolipideihinsä ja kalvoihinsa.
Siitä on sitten paikallisesti purettavissa takaisin esiin arakidonihappoa, jota taas entsyymisetti modifioi työkaluksi paikallisiin tehtäviin, immunologisiin, reaktiivisiin, tulehduksellisiin, signaloiviin ym tarpeista riippuen.
Koska arakidonihapossa ( eikosatetraeenihapossa) on 20 hiiltä (eicosa), niin sen johdannaisia sanotaan eikosanoideiksi.
Tetraeeni tarkoittaa että siinä on 4 kpl kaksoissidoksia rakenteessa.
Eri entsyymiseteistä sitten lähtee käyntiin eikosanoidikaskadit. Ne ovat myös kudosspesifisiä ( verisuonten puolella, keuhkoissa, ihossa, eri solukalvoissa, valkosoluissa, aivoissa jne on omat tyyppireaktionsa ja omat molekyylinsä).
PG= prostaglandiinit
LT= leukotrieenit
LX= lipoxygenaasit
HPETE=hydroperoxieikosatetraeenihappo
http://upload.wikimedia.org/wikipedia/commons/thumb/4/40/Eicosanoid_synthesis.svg/400px-Eicosanoid_synthesis.svg.png
Mitä kaikkea vapaasta arakidonihaposta voi tulla mm?
http://groundupstrength.wdfiles.com/local--files/health:omega-3-fatty-acids-and-inflammation/eicosanoid%20biosynthesis.JPG
Tämä on tosi aggressiivisten reaktiivisten molekyylien puu.
Ne kovasti mainostetut kalarasvat muodostavat näitten sukuisia molekyylejä ,jotka ovat vähemmän aggressiivisia ja täten vaimentavat ja miedontavat reaktiivisen arakidonihapon eikosanoidikaskadia.
Kts. Eikosanoidi keuhkofibroosissa
http://www.clinsci.org/cs/108/cs1080479.htm#CitedBy
lördag 11 december 2010
{kind=link}
{kind=link}
Aspiriini moduloi rasvahappoaineenvaihduntaa
http://scienceblog.cancerresearchuk.org/2010/12/07/aspirin-cuts-risk-of-dying-from-several-types-of-cancer/
JOS niin on että aspiriini on edullinen monia syövän kehittymisiä vastaan, niin mikä aiheuttaa syövän? Aspiriini jarruttaa arakidonihappoaineenvaihdunnan eikosanoidikaskadia, mutta ei sen kaikkia teitä.
http://jcp.bmj.com/content/57/1/6/F1.large.jpg
http://img.medscape.com/fullsize/migrated/405/963/ch1185.02.lieb.fig1.gif
Ei voi sanoa edes niin, että arakidonihapon vähempi saatavuus olisi edullisempaa, sillä sen normaali saatavuus pitää eikosanoidijärjestelmän kaikki säätelyt tasapainossa.
Arakidonihapon puute taas on hyvin paha tilanne, missä ihmisen solut eivät korjaannu. Arakidonihappoa asettuu kaikkiin solukalvoihin koko kehossa. Se on linolihappoperäistä.
Eikosandoi ja solukorjautumisjärjestelmän suistuminen tulehdukselliselle tielle ja tulehduksen jatkuminen nukleaariseen tasoon, solun muuttumiseen syöpäsoluksi, täytyy olla jokin rasvahappo ym aineenvaihdunnan poikkeusmolekyyli syynä eikä normaalin eikosanoidikaskadin kuvan kauniit molekyylit. MUTTA toisaalta, eikosanoidijärjestelmän työmolekyylien valmistuksessa tulee aika runsasti niitä sivurakenteita ja niistä käsin voi luonnollisesti löytyä karsinogeenejä, jos ne eivät häviä omissa silppureissaan.
JOS niin on että aspiriini on edullinen monia syövän kehittymisiä vastaan, niin mikä aiheuttaa syövän? Aspiriini jarruttaa arakidonihappoaineenvaihdunnan eikosanoidikaskadia, mutta ei sen kaikkia teitä.
http://jcp.bmj.com/content/57/1/6/F1.large.jpg
{kind=link}
http://img.medscape.com/fullsize/migrated/405/963/ch1185.02.lieb.fig1.gif
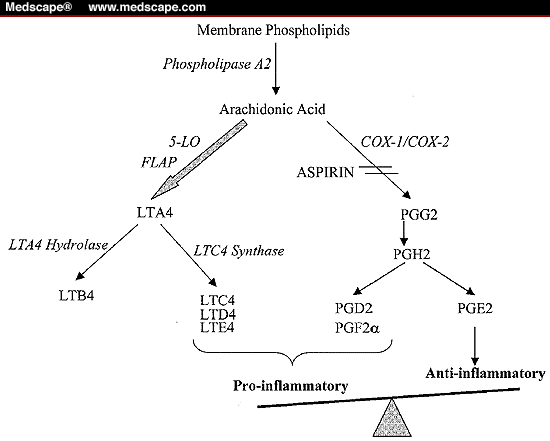{kind=link}
Ei voi sanoa edes niin, että arakidonihapon vähempi saatavuus olisi edullisempaa, sillä sen normaali saatavuus pitää eikosanoidijärjestelmän kaikki säätelyt tasapainossa.
Arakidonihapon puute taas on hyvin paha tilanne, missä ihmisen solut eivät korjaannu. Arakidonihappoa asettuu kaikkiin solukalvoihin koko kehossa. Se on linolihappoperäistä.
Eikosandoi ja solukorjautumisjärjestelmän suistuminen tulehdukselliselle tielle ja tulehduksen jatkuminen nukleaariseen tasoon, solun muuttumiseen syöpäsoluksi, täytyy olla jokin rasvahappo ym aineenvaihdunnan poikkeusmolekyyli syynä eikä normaalin eikosanoidikaskadin kuvan kauniit molekyylit. MUTTA toisaalta, eikosanoidijärjestelmän työmolekyylien valmistuksessa tulee aika runsasti niitä sivurakenteita ja niistä käsin voi luonnollisesti löytyä karsinogeenejä, jos ne eivät häviä omissa silppureissaan.
torsdag 4 november 2010
Endokannabinoidisysteemi endokriinisessä säätelyssä ja energiatasapainossa
Endocrine Reviews 27 (1): 73-100 The Emerging Role of the Endocannabinoid System in Endocrine Regulation and Energy Balance
Endocrinology Unit, Department of Internal Medicine and Gastroenterology et al.
During the last few years, the endocannabinoid system has emerged as a highly relevant topic in the scientific community. Endokannabinoidien nimiin on jo kirjattu monia erilaisia säätelytoimintoja ja niitten osuutta useissa patofysiologisissa tiloissa tutkitaan intensiivisesti. Many different regulatory actions have been attributed to endocannabinoids, and their involvement in several pathophysiological conditions is under intense scrutiny. Kannabinoidireseptorit CB1 ja CB2 löydettiin alunperin Cannabis sativa-nimisen kasvin psykotrooppisen komponentin ( cannabinoidien) reseptoreina. Nämä reseptorit osallistuvat monien keskushermoston ja perifeerisen hermoston fysiologisten funktioitten säätelyyn. Cannabinoid receptors, named CB1 receptor and CB2 receptor, first discovered as the molecular targets of the psychotropic component of the plant Cannabis sativa, participate in the physiological modulation of many central and peripheral functions. CB2 reseptoria esiintyy lähinnä immuunisoluissa, kun taas CB1 reseptori on runsain G-proteiiniin kytkeytynyt reseptori, mitä aivoissa ilmenee. CB2 receptor is mainly expressed in immune cells, whereas CB1 receptor is the most abundant G protein-coupled receptor expressed in the brain. CB1 reseptoria ilmenee hypotalamuksessa ja hypofyysissä. Tiedetään, että sen aktivaatio moduloi kaikkia endokriinisia hypotalamo-perifeerisiä endokriinisia akseleita. CB1 receptor is expressed in the hypothalamus and the pituitary gland, and its activation is known to modulate all the endocrine hypothalamic-peripheral endocrine axes. On lisääntyvää näyttöä, että tämä EC-järjestelmä omaa osuuden stressivasteessa vaikuttamalla hypotalamus-hypofyysi-lisämunuais-akseliin ja reproduktion kontrollissa modifioimalla gonadotropiinien vapautumista, fertiliteettiä ja seksuaalista käyttäytymistä. An increasing amount of data highlights the role of the system in the stress response by influencing the hypothalamic-pituitary-adrenal axis and in the control of reproduction by modifying gonadotropin release, fertility, and sexual behavior.Endokannabinoidijärjestelmän (EC) kyky kontrolloida ruokahalua, ravinnonottoa ja energiatasapainoa on saanut viime aikoina myös suurta huomiota erityisesti näiden funktioiden taustatekijöiden suhteen. The ability of the endocannabinoid system to control appetite, food intake, and energy balance has recently received great attention, particularly in the light of the different modes of action underlying these functions. Endokannabinoidijärjestelmä moduloi ravinnon palkitsevaisuutta vaikuttamalla erityisiin mesolimbisiin alueisiin aivoissa. The endocannabinoid system modulates rewarding properties of food by acting at specific mesolimbic areas in the brain. In the hypothalamus, CB1 receptor and endocannabinoids are integrated components of the networks controlling appetite and food intake. Mielenkiintoista on , että endokannabinoidien on viime aikoina osoitettu kontrolloivan aineenvaihdunnallisia funktioita perifeerisissä kudoksissa kuten adiposyyteissä, maksasoluissa, mahasuolikanavassa ja mahdollisesti tahdonalaisessa lihaksessa. Interestingly, the endocannabinoid system was recently shown to control metabolic functions by acting on peripheral tissues, such as adipocytes, hepatocytes, the gastrointestinal tract, and, possibly, skeletal muscle. Järjestelmän relevanssia vahvistaa edelleen huomio siitä, että ECV-järjestelmään interaktiossa olevien lääkkeiden on havaittu olevan lupaavia kandidaatteja erilaisten sairauksien hoitoon, joihin luetaan myös liikalihavuus. The relevance of the system is further strenghtened by the notion that drugs interfering with the activity of the endocannabinoid system are considered as promising candidates for the treatment of various diseases, including obesity. |
Anadamidi ja dopaminerginen neuroni
- Anadamidi ja DA neuroni
Tässä on kuva neuronista joka on dopaminerginen. Tämän kuvan laitoin pohdintoja varten. Tämä on hahmoteltu vuonna 1999.
- Anadamidihermotuksen kliinisestä merkityksestä. Pohdintaa varten (ADHD alue) Tässä tutkimuksessa oli nostettu EPA osuutta PUFA hapoissa ja vaste oli neurologisesti edullinen ADHD:ssa. Supplementit, joissa on korkea annos EPA/DHA konsentraatteja voivat parantaa ADHD lasten käytösongelmia. Tämä artikkeli ei suoraan mainitse anadamidin osuutta, mutta sillä on epäsuorasti merkittävä osuus, kunhan sen saa selville.
CONCLUSION: The findings of this small pilot study suggest supplementation with high-dose EPA/DHA concentrates may improve behavior in children with ADHD.
Effects of an open-label pilot study with high-dose EPA/DHA concentrates on plasma phospholipids and behavior in children with attention deficit hyperactivity disorder. Sorgi PJ, Hallowell EM, Hutchins HL, Sears B. Nutr J. 2007 Jul 13;6:16.PMID: 17629918 [PubMed - indexed for MEDLINE]Free PMC ArticleFree text
EPA ( C20:5 n6) and anandamide
J Nutr Biochem. 2010 Oct 14. Eicosapentaenoic acid decreases expression of anandamide synthesis enzyme and cannabinoid receptor 2 in osteoblast-like cells. Hutchins HL, Li Y, Hannon K, Watkins BA. Lipid Chemistry and Molecular Biology Laboratory, Purdue University, West Lafayette, IN 47907, USA.
Abstract (Suomennosta)
ANANDAMIDI (AEA) on endogeeninen agonisti CB2 reseptorille, jota ilmenee osteoblasteissa.
Anandamide (AEA) is an endogenous agonist for the cannabinoid receptor 2 (CB2) which is expressed in osteoblasts.
ARAKIDONIHAPPO (AA) on anandamidin prekursoriaine; ravintoperäisen n3- linjaisen PUFA rasvahapon tiedetään vähentävän arakidonihapon pitoisuutta kudoksissa ja soluissa.
Arachidonic acid (AA) is the precursor for AEA and dietary n-3 polyunsaturated fatty acids (PUFA) are known to reduce the concentrations of AA in tissues and cells.
- Hypoteesi
Tästä tutkijaryhmä teki hypoteesinsa siitä, että n3 PUFA hapot, eikosapentaeenihappo (EPA) ja dokosaheksaeenihappo (DHA), jotka vähentävät solujen arakidonihappopitoisuuksia, voisivat alentaa anandamidin pitoisuutta osteoblasteissa muuntamalla endokannabinoidisysteemin entsyymien ilmenemistä.
Therefore, we hypothesized that n-3 PUFA, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which reduce AA in cells, could lower AEA in osteoblasts by altering enzyme expression of the endocannabinoid (EC) system.
- (Tekniikasta selitystä:
MC3T3-E1 osteoblast-like cells were grown for 6, 10, 15, 20, 25 or 30 days in osteogenic medium. Osteoblasts were treated with 10 μM of AA, EPA, DHA, oleic acid (OA) or EPA+DHA (5 μM each) for 72 h prior to their collection for measurement of mRNA and alkaline phosphatase (ALP) activity.)
- PUFA n6 ja n3 linjoista
Osteoblastit, joita käsiteltiin arakidonihapolla sisälsivät enemmän arakidonihappoa ja n6- linjan PUFA happoja kuin ne joita käsiteltiin EPA ja DHA-hapoilla, joilla oli matalampi n6 linjan kuin n3- linjan happojen pitoisuus
Compared to vehicle control, osteoblasts treated with AA had higher levels of AA and n-6 PUFA while those treated with EPA and DHA had lower n-6 but higher n-3 PUFA.
- Tuloksista
Riippumatta rasvahappokäsittelystä osteoblastit kypsyivät normaalisti, mikä ilmeni ALP-entsyymistä.
Independent of the fatty acid treatments, osteoblasts matured normally as evidenced by ALP activity.
- NAPE-PLD, FAAH, CB2
NAPE-selektiivisen PLD-entsyymin, rasvahappoamidihydrolaasin (FAAH) ja CB2mRNA:n ilmenemät olivat suuremmat 20 päivän kuin 10 päivänä tuloksissa.
N-acyl phosphatidylethanolamine-selective phospholipase D (NAPE-PLD), fatty acid amide hydrolase (FAAH) and CB2 mRNA expression were higher at 20 days compared to 10 days.
NAPE-PLD ja CB2mRNA ilmenemät olivat matalammat niissä osteoblasteissa, joita oli käsitelty EPA rasvahapolla verrattuna kaikkiin muihin ryhmiin.
NAPE-PLD and CB2 mRNA was lower in osteoblasts treated with EPA compared to all other groups.
Täten havaitaan, että NAPE-PLD mRNA, FAAH mRNA ja CB2mRNA lisääntyivät osteoblastin kypsymisen aikana ja että EPA rasvahappo vähensi NAPE-PLD entsyymin ja CB2 reseptorin mRNA:ta
Thus, mRNA expression for NAPE-PLD, FAAH, and CB2 increased during osteoblast maturation and EPA reduced mRNA for NAPE-PLD and CB2 receptor.
- Yhteenveto
EPA alensi endokannabinoidisysteemin(EC) proteiinien mRNA pitoisuuksia ja anandamidiaineenvaíhdunnan synteesivaiheen ja hajoamisvaiheen aineenvaihdunnan entsyymien mRNA-pitoisuuksia osteoblasteissa.
In conclusion, EPA lowered mRNA levels for proteins of the EC system and mRNA for AEA synthesis/degradation is reported in osteoblasts.
LinkOut - more resources
söndag 31 oktober 2010
FOSFOLIPIDI
Solujen kalvot ovat fosfolipidien kaksoiskerroksia.
Fosfolipidi (PL) sijoittaa glyserolirakneteiseen hyllymäiseen sijoitukseen tärkeitä rasvahappoja, joita tarvitaessa ( on demand) hyödynnetään eri puolilla kehoa. Yksi näistä kaikkiin soluihin sijoitetusta rasvahaposta on arakidonihappo (AA), jota on saatavilla tarpeen mukaan ja se osallistuu kehon sorvauksiin ja plastisuuden ylläpitämisiin.
Types of phospholipid, fosfolipidityypit
Diacylglyceride structures, diglyseridirakenteet
Phosphosphingolipids, Fosfosfingolipidit
Phospholipid derivatives, fosfolipidijohdannaiset
On geneettisesti ohjautunutta mihin asemaan minkäkinlainen rasvahappo mieluiten asettuu , essentielleillä rasvahapoilla (EFA) on erityinen prioriteettinsa. Jos niitä ei ole valiutuu non-essentiellejä näille paikoille.
Essentiellit rasvahapot linolihappo ja linoleenihappo pystyvät kehoentsyymien kautta muodostamaan sellaisia rasvahappoja, joihin kehon korkea-asteisesti spesifiset entsyymijärjestelmät voivat pureutua ja käyttää näitä molekyyleja tarkoin säädeltyinä työaseina kehon hoidossa, kuten esim avaruusasemaa hoidettaessa, kun korjataan jotain modulia ulkoavaruuden puolella ja käytetään jotain työasetta kädessä.
Arakidonihappojärjestelmä on tällainen huipputäsmällinen työase kehon kunnossapidossa. Se otetaan esiin näistä rakenteellisista säilöpaikosita rakenteellisista lipideistä ja muokataan soveltuvaksi työaseeksi.
Tällainen työase otetaan vain tarvittaessa esiin, "on demand" ja hoidetaan sitten inaktiiviseen muotoon takaisin työn jälkeen tavalla tai toisella.
Koska linolihapoista muodostuva ARAKIDONIHAPPO on aika aggressiivinen aine sen johdannaisten muodostuttua, linoleenihaposta käsin on sitä järejstelmäkin olemassa paralleelisti. sofistisesti vaimentavaLinoleenihappolinjasta tunnetaan EPA ja DHA joita voi saada vlamiissakin muodossa kalarasvoista käsin.
Fosfolipidi (PL) sijoittaa glyserolirakneteiseen hyllymäiseen sijoitukseen tärkeitä rasvahappoja, joita tarvitaessa ( on demand) hyödynnetään eri puolilla kehoa. Yksi näistä kaikkiin soluihin sijoitetusta rasvahaposta on arakidonihappo (AA), jota on saatavilla tarpeen mukaan ja se osallistuu kehon sorvauksiin ja plastisuuden ylläpitämisiin.
Types of phospholipid, fosfolipidityypit
Diacylglyceride structures, diglyseridirakenteet
- See: Glycerophospholipid
- Phosphatidic acid (phosphatidate) (PA)
- Phosphatidylethanolamine (cephalin) (PE)
- Phosphatidylcholine (lecithin) (PC)
- Phosphatidylserine (PS)
- Phosphoinositides:
- Phosphatidylinositol (PI)
- Phosphatidylinositol phosphate (PIP)
- Phosphatidylinositol bisphosphate (PIP2) and
- Phosphatidylinositol triphosphate (PIP3).
Phosphosphingolipids, Fosfosfingolipidit
- Ceramide phosphorylcholine (Sphingomyelin) (SPH)
- Ceramide phosphorylethanolamine (Cer-PE)
- Ceramide phosphorylglycerol
Phospholipid derivatives, fosfolipidijohdannaiset
- See table below for an extensive list.
- Natural phospholipid derivates:
- egg PC, egg PG, soy PC, hydrogenated soy PC, sphingomyelin as natural phospholipids.
- Synthetic phospholipid derivates:
- Phosphatidic acid (DMPA, DPPA, DSPA)
- Phosphatidylcholine (DDPC, DLPC, DMPC, DPPC, DSPC, DOPC, POPC, DEPC)
- Phosphatidylglycerol (DMPG, DPPG, DSPG, POPG)
- Phosphatidylethanolamine (DMPE, DPPE, DSPE DOPE)
- Phosphatidylserine (DOPS)
- PEG phospholipid (mPEG-phospholipid, polyglycerin-phospholipid, funcitionalized-phospholipid, terminal activated-phospholipid)
Abbreviations used and chemical information of glycerophospholipids
Abbreviation | CAS | Name | Type |
|---|---|---|---|
| DDPC | 3436-44-0 | 1,2-Didecanoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DEPA-NA | 80724-31-8 | 1,2-Dierucoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DEPC | 56649-39-9 | 1,2-Dierucoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DEPE | 988-07-2 | 1,2-Dierucoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DEPG-NA | 1,2-Dierucoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol | |
| DLOPC | 998-06-1 | 1,2-Dilinoleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DLPA-NA | 1,2-Dilauroyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid | |
| DLPC | 18194-25-7 | 1,2-Dilauroyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DLPE | 1,2-Dilauroyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine | |
| DLPG-NA | 1,2-Dilauroyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol | |
| DLPG-NH4 | 1,2-Dilauroyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol | |
| DLPS-NA | 1,2-Dilauroyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DMPA-NA | 80724-3 | 1,2-Dimyristoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DMPC | 18194-24-6 | 1,2-Dimyristoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DMPE | 988-07-2 | 1,2-Dimyristoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DMPG-NA | 67232-80-8 | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DMPG-NH4 | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol | |
| DMPG-NH4/NA | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium/Ammonium Salt) | Phosphatidylglycerol | |
| DMPS-NA | 1,2-Dimyristoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DOPA-NA | 1,2-Dioleoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid | |
| DOPC | 4235-95-4 | 1,2-Dioleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DOPE | 4004-5-1- | 1,2-Dioleoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DOPG-NA | 62700-69-0 | 1,2-Dioleoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DOPS-NA | 70614-14-1 | 1,2-Dioleoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine |
| DPPA-NA | 71065-87-7 | 1,2-Dipalmitoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DPPC | 63-89-8 | 1,2-Dipalmitoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DPPE | 923-61-5 | 1,2-Dipalmitoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DPPG-NA | 67232-81-9 | 1,2-Dipalmitoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DPPG-NH4 | 73548-70-6 | 1,2-Dipalmitoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol |
| DPPS-NA | 1,2-Dipalmitoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DSPA-NA | 108321-18-2 | 1,2-Distearoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DSPC | 816-94-4 | 1,2-Distearoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DSPE | 1069-79-0 | 1,2-Distearoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DSPG-NA | 67232-82-0 | 1,2-Distearoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DSPG-NH4 | 108347-80-4 | 1,2-Distearoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol |
| DSPS-NA | 1,2-Distearoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| Egg Sphingomyelin empty Liposome | |||
| EPC | Egg-PC | Phosphatidylcholine | |
| HEPC | Hydrogenated Egg PC | Phosphatidylcholine | |
| HSPC | High purity Hydrogenated Soy PC | Phosphatidylcholine | |
| HSPC | Hydrogenated Soy PC | Phosphatidylcholine | |
| LYSOPC MYRISTIC | 18194-24-6 | 1-Myristoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| LYSOPC PALMITIC | 17364-16-8 | 1-Palmitoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| LYSOPC STEARIC | 19420-57-6 | 1-Stearoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| Milk Sphingomyelin MPPC | 1-Myristoyl-2-palmitoyl-sn-glycero 3-phosphocholine | Phosphatidylcholine | |
| MSPC | 1-Myristoyl-2-stearoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| PMPC | 1-Palmitoyl-2-myristoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| POPC | 26853-31-6 | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| POPE | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine | |
| POPG-NA | 81490-05-3 | 1-Palmitoyl-2-oleoyl-sn-glycero-3[Phospho-rac-(1-glycerol)...] (Sodium Salt) | Phosphatidylglycerol |
| PSPC | 1-Palmitoyl-2-stearoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| SMPC | 1-Stearoyl-2-myristoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| SOPC | 1-Stearoyl-2-oleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine | |
| SPPC | 1-Stearoyl-2-palmitoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
- PC tarkoittaa fosfatidyylikoliineja, lesitiinejä
- PS tarkoittaa fosfatidyyliseriinejä
- PE tarkoittaa fosfatidyylietanolamineja, kefaliineja
- PI tarkoittaa lipositoleja, fosfoinositolilajeja
- LPA tarkoittaa lysolesitiiniä, lysofsofatidyylikoliineja
- PA tarkoittaa fosfatidihappoa
- Ptd-glyserolikin on listoissa
On geneettisesti ohjautunutta mihin asemaan minkäkinlainen rasvahappo mieluiten asettuu , essentielleillä rasvahapoilla (EFA) on erityinen prioriteettinsa. Jos niitä ei ole valiutuu non-essentiellejä näille paikoille.
Essentiellit rasvahapot linolihappo ja linoleenihappo pystyvät kehoentsyymien kautta muodostamaan sellaisia rasvahappoja, joihin kehon korkea-asteisesti spesifiset entsyymijärjestelmät voivat pureutua ja käyttää näitä molekyyleja tarkoin säädeltyinä työaseina kehon hoidossa, kuten esim avaruusasemaa hoidettaessa, kun korjataan jotain modulia ulkoavaruuden puolella ja käytetään jotain työasetta kädessä.
Arakidonihappojärjestelmä on tällainen huipputäsmällinen työase kehon kunnossapidossa. Se otetaan esiin näistä rakenteellisista säilöpaikosita rakenteellisista lipideistä ja muokataan soveltuvaksi työaseeksi.
Tällainen työase otetaan vain tarvittaessa esiin, "on demand" ja hoidetaan sitten inaktiiviseen muotoon takaisin työn jälkeen tavalla tai toisella.
Koska linolihapoista muodostuva ARAKIDONIHAPPO on aika aggressiivinen aine sen johdannaisten muodostuttua, linoleenihaposta käsin on sitä järejstelmäkin olemassa paralleelisti. sofistisesti vaimentavaLinoleenihappolinjasta tunnetaan EPA ja DHA joita voi saada vlamiissakin muodossa kalarasvoista käsin.
Endokannabinoiditiedemiehet
Sign-in free and Explore the Exciting World of BiomedExperts:
- Over 1,800,000 Profiles
- More than 3,500 Organizations worldwide
- State of the Art Network Visualizations
- Manage your own Profile
- Locate Experts in your Country/Region
- Locate Experts in your 1. and 2. Level Network
- Connect to Experts Worldwide
NetworkView

Experts in Endocannabinoids
Endogenous compounds that bind to CANNABINOID RECEPTORS. They are structurally different from the CANNABINOIDS which are primarily derived from plants.
lördag 30 oktober 2010
Anandamidi. Endokannabinoidien biosynteesistä
Anandamidin tehtävistä ja muodostuksesta
Aktiviteetti havaittiin suurimmaksi paikallisesti ajatellen hippokampissa, sitten thalamuksessa, cortex alueessa ja striatumissa.
Entsyymin aktiivisuus oli matalinta pikkuaivoissa, aivosillassa (pons) ja ydinjatkoksessa ( medulla oblongata)
The ability of brain tissue to enzymatically synthetize anandamide and the existence of specific receptors for this eicosanoid suggest the presence of anadamide containing anandaergic neurons.
Koska aivot pystyvät syntetisoimaan entsymaattisesti anadamidia ja sille on olemassa omat reseptorinsa, asia viittaa siihen, että on olemassa andandamidia sisältäviä anandaergisiä hermoja
Lähde: W A Devane et J Axelrod ( PNAS 1994)
HUOM.
Toivottavasti lukija ymmärtää, että huumemolekyyli marihuana, cannabis, THC on toksiini, joka asettuu anandaergiseen järjestelmään ja estää tämän normaalin synaptisen plastisuuden . Lisäksi THC on rasvaliukoinen ja jää pitkäksi aikaa aivoon, kuukausiksi, joten anandaerginen järjestelmä voi lopulta irreversibelisti jäädä pois toiminnasta myrkyttyneenä.
Toivottavasti artikkelisarjani vaikuttaa että ihmiset alkavat lopettaa marihuanan ym sen sukuisten myrkkyjen lataamisen aivojensa rasvamolekyyliin ja alkavat käyttää kunnon ruokaa, puhtaita kasvis- ja kalaöljyjä ja muuta normaalia ruokaa, jotta tämä järjestelmä voisi elpyä ja ehkä pystyä oksidoimaan jollain tavalla kellan vihreäksi aivon värjääviä THC molekyylejä vuosien kuluessa ja saada taas normaalin vitivalkoisen valkoharmaan aivonsa takaisin.
- AEA on lipiditransmittoriaine jota syntetisoituu tarpeen mukaan ( on demand) aivoissa neuroneista eikä sitä varastoidu mitenkään rakkuloihin kuten tavallisia hermonvälittäjäaineita.
- Kuten Wikipedian tekstistä käy ilmi, sitä muodostuu seurauksena aktiiville neuronitoiminnan sekventiaaliselle tapahtumalle, jolloin se muodostaa tarpeen mukaan plastista materiaalia synapsitienoossa ja vaikuttaa neuronin fysikokemiallista sopeutumista aktiivisuuteensa ja impulssinkuljetustehtäväänsä. Siis käyttämällä aktiivisti aivojaan pitää aivojaan kunnossa samalla tällä luonnonjärjestelmällä.
- Myös makrofagit kehittävät anandamidia (AÈA) ja niissä indusoi mm. toksinen LPS anandamidin syntetisoitumisen. AEA synteesiä on nähty syynä septisen shokin hypotensiiviseen reaktioon ja maksakirroosin vakavaan muotoon.
- Anandamidi (AEA) voi syntetisoitua kalvoissa esiintyvästä edeltäjämolekyylistä N-arachidonoyl phosphatidylethanolamine (NAPE) siten, että se pilkkoutuu fosfolipaasientsyymillä NAPE-PLD
- Makrofagissa katalysoi PLC entsyymi NAPE- molekyyliä ja sitä sanotaan NAPE-PLC entsyymiksi. Muodostuu fosfoanandamidia ( pAEA), joka defosforyloituu mm PTPN22 :lla ja antaa lopulta anandamidia (AEA),
- Jos LPS indusoi anandamidin muodostusta, säätyy NAPE-PLC ylös ja NAPE-PLD alas.
- Autoimmuunitaudeissa vallitsee anandamidin synteesissä PLC-fosfataasitie.
- Anadamidisynthase- entsyymin lokalisaatiosta
Aktiviteetti havaittiin suurimmaksi paikallisesti ajatellen hippokampissa, sitten thalamuksessa, cortex alueessa ja striatumissa.
Entsyymin aktiivisuus oli matalinta pikkuaivoissa, aivosillassa (pons) ja ydinjatkoksessa ( medulla oblongata)
The ability of brain tissue to enzymatically synthetize anandamide and the existence of specific receptors for this eicosanoid suggest the presence of anadamide containing anandaergic neurons.
Koska aivot pystyvät syntetisoimaan entsymaattisesti anadamidia ja sille on olemassa omat reseptorinsa, asia viittaa siihen, että on olemassa andandamidia sisältäviä anandaergisiä hermoja
Lähde: W A Devane et J Axelrod ( PNAS 1994)
HUOM.
Toivottavasti lukija ymmärtää, että huumemolekyyli marihuana, cannabis, THC on toksiini, joka asettuu anandaergiseen järjestelmään ja estää tämän normaalin synaptisen plastisuuden . Lisäksi THC on rasvaliukoinen ja jää pitkäksi aikaa aivoon, kuukausiksi, joten anandaerginen järjestelmä voi lopulta irreversibelisti jäädä pois toiminnasta myrkyttyneenä.
Toivottavasti artikkelisarjani vaikuttaa että ihmiset alkavat lopettaa marihuanan ym sen sukuisten myrkkyjen lataamisen aivojensa rasvamolekyyliin ja alkavat käyttää kunnon ruokaa, puhtaita kasvis- ja kalaöljyjä ja muuta normaalia ruokaa, jotta tämä järjestelmä voisi elpyä ja ehkä pystyä oksidoimaan jollain tavalla kellan vihreäksi aivon värjääviä THC molekyylejä vuosien kuluessa ja saada taas normaalin vitivalkoisen valkoharmaan aivonsa takaisin.
Endokannabinoidien oksidaatioteistä
Natasha T Snider, Vyvyca J Walker, P F Hollenberg
Oxidation of the endogenous cannabinoid arachidonoyl ethanolamide (AEA) by the cytochrome P450 monooxygenases: Physiological and pharmacological implications.
Anandamidin (AEA) pääasiallinen inaktivoiva entsyymi on FAAH, rasvahappoamidihydrolaasi.
Tämän molekyylin, kehon oman endokannabinoidin anandamidin (AEA) oksidaatio tapahtuu ihmisessä monella P450 entsyymillä kuten CYP3A4, CYP4F2, CYP4X1 entsyymeillä ja hyvin polymorfisella CYP2D6 entsyymillä ja näin muodostuu lukuisia rakenteellisesti erilaisia lipidejä, joilla todennäköisesti on tärkeitä fysiologisia tehtäviä kuten esimerkiksi on osoitettu siitä epoksidista mikä anadamidijohdannaisena muodostuu, Se on voimakas CB2 reseptorin agonisti, myötävaikuttaja.
Tässä artikkelissa koetetaan painottaa CB2 reseptorin osuutta ja sitä miten tärkeää on ymmärtää anandamidin (AEA) aineenvaihdunnan P450-välitteiset tiet, koska ne ovat todennäköisesti tärkeitä endokannabinoidien signaloinnin välittämisessä ja myös endokannabinoideihin kohdistettujen lääkkeitten farmakologisisssa vasteissa.
20-HEET-EA , 20-hydroxyepoxyeicosatrienoic acid ethanol amide
20-HETE , 20 hydroxyeicosatetraenoic acid
20-HETE-EA, 20- hydroxyeicosatetraenoic acid ethanolamide
2AG, 2 arachidonoylglycerol (endocannabinoidi laji)
ABHD4, alfa/beta-hydrolase 4
CB1, canabinoidireceptor 1
CB2 , cannabinoidrecepotor 2
CNS, Central Nervous System
COX, Cyclooxygenase
DHET, dihydroxyeicosatrienoic acid
EA, ethanolamide
EG, epoxyeicosatrienoylglycerol
EET, epoxyeicosatrienoicacid
FAAH, fattyacid hydrolylase
FABP, fattyacid binding protein
CDE1, glycerofosfodiesterase
GP, glycerofosfo
HSC, hepatic stellate cell
IFNgamma, interferone gamma
LOX, lipoxygenase
LPS
NAE, N-acylethanolamine
NAPE, N-arachidonoylphosphatidylethanolamine
P450
PLD, phospholipase D
PPAR, peroxisome proliferator activated receptor
PTPN22, protein tyrosine phosphatase
SNL, spinal nerve- ligated
sPLA2, soluble PLA2
TRPV1 transietn receptorpotential vanilloind type-1 receptor
TRPV4, transient receptor potential channel 4
Oxidation of the endogenous cannabinoid arachidonoyl ethanolamide (AEA) by the cytochrome P450 monooxygenases: Physiological and pharmacological implications.
Anandamidin (AEA) pääasiallinen inaktivoiva entsyymi on FAAH, rasvahappoamidihydrolaasi.
Tämän molekyylin, kehon oman endokannabinoidin anandamidin (AEA) oksidaatio tapahtuu ihmisessä monella P450 entsyymillä kuten CYP3A4, CYP4F2, CYP4X1 entsyymeillä ja hyvin polymorfisella CYP2D6 entsyymillä ja näin muodostuu lukuisia rakenteellisesti erilaisia lipidejä, joilla todennäköisesti on tärkeitä fysiologisia tehtäviä kuten esimerkiksi on osoitettu siitä epoksidista mikä anadamidijohdannaisena muodostuu, Se on voimakas CB2 reseptorin agonisti, myötävaikuttaja.
Tässä artikkelissa koetetaan painottaa CB2 reseptorin osuutta ja sitä miten tärkeää on ymmärtää anandamidin (AEA) aineenvaihdunnan P450-välitteiset tiet, koska ne ovat todennäköisesti tärkeitä endokannabinoidien signaloinnin välittämisessä ja myös endokannabinoideihin kohdistettujen lääkkeitten farmakologisisssa vasteissa.
- Tässä alueessa tarvittavaa sanastoa
20-HEET-EA , 20-hydroxyepoxyeicosatrienoic acid ethanol amide
20-HETE , 20 hydroxyeicosatetraenoic acid
20-HETE-EA, 20- hydroxyeicosatetraenoic acid ethanolamide
2AG, 2 arachidonoylglycerol (endocannabinoidi laji)
ABHD4, alfa/beta-hydrolase 4
CB1, canabinoidireceptor 1
CB2 , cannabinoidrecepotor 2
CNS, Central Nervous System
COX, Cyclooxygenase
DHET, dihydroxyeicosatrienoic acid
EA, ethanolamide
EG, epoxyeicosatrienoylglycerol
EET, epoxyeicosatrienoicacid
FAAH, fattyacid hydrolylase
FABP, fattyacid binding protein
CDE1, glycerofosfodiesterase
GP, glycerofosfo
HSC, hepatic stellate cell
IFNgamma, interferone gamma
LOX, lipoxygenase
LPS
NAE, N-acylethanolamine
NAPE, N-arachidonoylphosphatidylethanolamine
P450
PLD, phospholipase D
PPAR, peroxisome proliferator activated receptor
PTPN22, protein tyrosine phosphatase
SNL, spinal nerve- ligated
sPLA2, soluble PLA2
TRPV1 transietn receptorpotential vanilloind type-1 receptor
TRPV4, transient receptor potential channel 4
CB2 endokannabinoidireseptorin merkitys
Diabetologia. 2010 Sep 11. [Epub ahead of print] Deficiency of CB2 cannabinoid receptor in mice improves insulin sensitivity but increases food intake and obesity with age. Agudo J, Martin M, Roca C, Molas M, Bura AS, Zimmer A, Bosch F, Maldonado R. Center of Animal Biotechnology and Gene Therapy, Department of Biochemistry and Molecular Biology, School of Veterinary Medicine, Universitat Autònoma de Barcelona, Bellaterra, Spain.
Abstract (suomennosta)
Endokannabinoidijärjestelmällä oletataan olevan avainmerkitystä energian varastoimisessa ja aineenvaihdunnallisissa häiriöissä. Endokannabinoidireseptori CB2R havaittiin ensiksi immuunisoluista. Sitä esiintyy perifeerisissä pääelimissä vastaamassa aineenvaihdunnan kontrollista.AIMS/HYPOTHESIS: The endocannabinoid system has a key role in energy storage and metabolic disorders. The endocannabinoid receptor 2 (CB2R), which was first detected in immune cells, is present in the main peripheral organs responsible for metabolic control.
Lihavuudessa CB2R osallistuu rasvakudoksen tulehduksen kehittymiseen ja rasvamaksan muodostukseen.
During obesity, CB2R is involved in the development of adipose tissue inflammation and fatty liver.
Tutkijat selvittivät CB2R puutteen vaikutuksia verensokerin, GLUKOOSIN aineenvaihduntaan
TULOKSISTA: CB2R puutteiselle hiirelle kehittyi suurempi ravinnon otto ja kehon paino
However, 12-month-old obese Cb2 (-/-) mice did not develop insulin resistance and showed enhanced insulin-stimulated glucose uptake in skeletal muscle.
Kuitenkaan 12 kk ikäiselle CB2- reseptorivajeiselle hiirelle ei kehittynyt insuliiniresistenssiä ja sillä esiintyi kohonnut insuliinin stimuloima glukoosin otto lihassoluun.
In agreement, adipose tissue hypertrophy was not associated with inflammation.
Saman suuntainen tulos saatiin, kun normaalille hiirelle annettiin CB2-reseptoriantagonistia. Sen insuliiniherkkyys parani.
Similarly, treatment of wild-type mice with CB2R antagonist resulted in improved insulin sensitivity.
Lisäksi jos 2 kk ikäiselle CB2-vajeiselle hiirelle annettiin rasvaista ravintoa, sen painonkehittyminen aleni ja sillä todettiin normaalia insuliiniherkkyyttä.
Moreover, when 2-month-old Cb2 (-/-) mice were fed a high-fat diet, reduced body weight gain and normal insulin sensitivity were observed.
YHTEENVETO ja TULOSTEN TULKINTA:
CB2R välitteisten vasteitten puuttuminen suojasi hiirtä sekä iästä riippuvaa että dieetistä johtuvaa insuliiniresistenssiä vastaan, mikä viittaisi siihen, että näissä reseptoreissa voi olla jotain terapeuttisen kohteen potentiaalia, jos on kyse lihavuudesta tai insuliiniresistenssistä.
CONCLUSIONS/INTERPRETATION: These results indicate that the lack of CB2R-mediated responses protected mice from both age-related and diet-induced insulin resistance, suggesting that these receptors may be a potential therapeutic target in obesity and insulin resistance.
KOMMENTTI:
Tässä ei voi vetää mitään suoraa yhteenvetoa siitä, että sammuttamalla CB2 reseptori jollain molekyylillä suoraan saataisiin aikaan etsittyjä edullisia vaikutuksia. Tässä täytyy ensin kaavoittaa koko CB-reseptorijärjestelmän luonne, miten se toimii, mikä sen aktivoi ja miten se säätyy. Otan näistä joitain detaljeja esiin, jotta voisi välttää bias harhateitä.
Tässä täytyy menna takaisin aivan arakidonihappoon ja anandamidiin asti.
fredag 29 oktober 2010
Endokannabinoidien sitoutuminen ja signaalinjohtuminen
Endocannabinoid binding & signal transduction
While there have been some papers that have linked concurrent stimulation of dopamine and CB1 receptors to an acute rise in cAMP production, it is accepted that CB1 activation causes an inhibition of cyclic adenosine monophosphate (or cAMP) when activated alone.
Tällaista cAMP inhibitiota seuraa fosforylaatio ja aktivaatio MAP-kinaasissa sekä PI3/PKB ja MEK/ERK teissä.
This inhibition of cAMP is followed by phosphorylation and subsequent activation of not only a suite (p38/p42/p44) of MAP kinases but also the PI3/PKB and MEK/ERK pathway (Galve-Roperh et al., 2002; Davis et al., 2005; Jones et al., 2005; Graham et al., 2006).
Sitten on tuloksia THC:n vaikutuksesta hippokampaalisiin geeneihin: Myeliinin baasisen proteiinin pitoisuus nousee. endoplasmisia proteiineja, sytokromipoksidaasia ja kahta adheesiomolekyyliä NCAM ja SC1 ilmenee. Alenemia puolestaan on kalmoduliinissa ja ribosomaalisessa RNA:ssa.
Results from rat hippocampal gene chip data after acute administration of tetrahydrocannabinol showed an increase in the expression of myelin basic protein, endoplasmic proteins, cytochrome oxidase, and two cell adhesion molecules: NCAM, and SC1; decreases in expression were seen in both calmodulin and ribosomal RNAs (Kittler et al., 2000).
Lisäksi CB1 aktivaation on osoitettu lisäävän transkriptiotekijöitten c-Fos ja Krox-24 aktiivisuutta.
In addition, CB1 activation has been demonstrated to increase the activity of transcription factors like c-Fos and Krox-24 (Graham et al., 2006).
Endocannabinoid binding & alterations in neuronal excitability
CB1-välitteisten muutosten molekulaarista mekanismia on myös tutkittu yksityiskohtaisesti kalvojännitteen vaihteluitten suhteen.
The molecular mechanisms of CB1-mediated changes to the membrane voltage have also been studied in detail.
CB1 myötävaikuttajat alentavat kalsiumin sisäänvirtausta blokeeraamalla jännitteestä riippuvien N, P/Q- ja L-tyyppisten kalsiumkanavien aktiivisuuden.
CB1 agonists reduce calcium influx by blocking the activity of voltage-dependent N-, P/Q- and L-type calcium channels.[10][11]
Lisäksi ne moduloivat kalsiumkanavien aktiivisuutta vaikuttaen G-proteiiniin kytkeytyneitten reseptorien alayksikköihin G1/0 ja Gs
In addition to acting on calcium channels, Gi/o and Gs, subunits of G protein-coupled receptors, activation has also been shown to modulate potassium channel activity.
Äskettäiset tutkimukset ovat osoittaneet, että CB1 aktivaatio kiihdyttää GIRK kaliumkanavaa, joka kuuluu Kir2- jonikanavaryhmään.
Recent studies have found that CB1 activation facilitates GIRK, a potassium channel belonging to the Kir3 family.[11] .
Tutkijat havaitsivat, että CB1 reseptori sijoittuu samaan paikkaan kuin kaliumkanavat GIRK ja Kv1.4 ja tästä , että näillä kahdella lienee keskinäistä vuorovaikutusta fysiologisissa puitteissa.
Corroborating Guo and Ikeda, Binzen et al. performed a series of immunohistochemistry experiments that demonstrated CB1 co-localized with GIRK and Kv1.4 potassium channels, suggesting that these two may interact in physiological contexts[12].
Keskushermostojärjestelmässä CB1 reseptorit enimmäkseen vaikuttavat epäsuorasti hermon ärtyvyyteen alentamalla tulevan synaptisen impulssivirran merkitystä.
In the central nervous system, CB1 receptors, for the most part, influence neuronal excitability indirectly, by reducing the impact of incoming synaptic input[13].
Tämän presynaptisen estämismekanismin uskotaan tapahtuvan silloin , kun neuroni postsynaptisesti vapauttaa endokannabinoideja retrogradisella, takaperoisella, tavalla ja ne kiinnittyvät CB1-reseptoreihin, joita esiintyy presynaptisen eli input neuronin hermoterminaalissa.
This mechanism ("presynaptic inhibition") is believed to occur when a neuron ("postsynaptic") releases endocannabinoids in a retrograde fashion, binding to CB1 receptors expressed on nerve terminals of an input neuron ("presynaptic").
Sitten CB1-reseptorit alentavat vapautuvien hermonvälittäjäaineitten pitoisuuksia, jolloin sen jälkeen tuleva input- signaalivirta presynaptisesta neuronista käsin ei ole enää niin vaikuttava postsynaptisessa neuronissa päin.
CB1 receptors then reduce the amount of neurotransmitter released, so that subsequent input from the presynaptic neuron has less of an impact on the postsynaptic neuron.
On todennäköistä, että presynaptinen inhibitio käyttää monia yllämainituista jonikanavamekanismeista, vaikka tuoretta näyttöä on siitä, että CB1 reseptorit voivat myös säätää hermonvälittäjäaineitten vapautumista mekanismilla, joka ei ole jonikanavaperäinen, kuten esim. AC-entsyymin ja PKA:n Gi/0 välitteisellä estämisellä.
It is likely that presynaptic inhibition uses many of the same ion channel mechanisms listed above, although recent evidence has shown that CB1 receptors can also regulate neurotransmitter release by a non-ion channel mechanism, i.e. through Gi/o mediated inhibition of adenylyl cyclase and Protein Kinase A[14]
On raportoitu myös CB1-reseptoreitten suorista vaikutuksista kalvoärtyvyyteen ja vahvasta vaikutuksesta kortikaalisten neuronien impulssinlähettämisiin
Still, direct effects of CB1 receptors on membrane excitability have been reported, and strongly impact the firing of cortical neurons[15]
On tehty koko joukko behavioristisia kokeita ja voitu osoittaa, että NMDA ( jonotrooppinen glutamaattireseptori) ja mGluR ( metabotrooppiset glutamaattireseptorit) toimivat CB1-reseptorin kanssa yhteisvaikutuksessa saaden aikaan hiirten analgesiaa ( kivuttomuutta) vaikka tämän vaikutuksen taustalla olevat seikät ovat selvittämättä.
In a series of behavioral experiments, Palazzo et al. demonstrated that NMDA, an ionotropic glutamate receptor, and the metabotropic glutamate receptors (mGluRs) work in concert with CB1 to induce analgesia in mice, although the mechanism underlying this effect is unclear.
Kaikenkaikkiaan näistä löydöistä käy ilmi, että CB1reseptori vaikuttaa neuronaaliseen ärtyvyyteen monilla mekanismeilla ja vaikutukset ovat relevantteja ja ilmenevät merkitsevällä tavalla havainto- ja käsityskyvyssä ja käyttäytymisessä
Together, these findings suggest that CB1 influences neuronal excitability by a variety of mechanisms, and these effects are relevant to perception and behavior.
KOMMENTTI:
Ihmisen terve havainto-ja käsityskyky perustuu aivoston hyvään kuntoon, synaptiseen plastisuuteen ja hyviin turn over- prosesseihin. Tämä järjestelmä perustuu arakidonihapon kautta tulevaan kykyyn uudistaa ja moduloida kaikkia kehon soluja, eri kohdissa kehoa eri tavalla, mutta aivoissa endokannabinoidijärjestelmällä (mikä on saanut hankalan nimen vaikka sen pitäisi olla nimi joka viittaa fysiologiseen kantasanaan, eikä järjestelmää myrkyttävään sanaan).
Anandamidijärjestelmällä on nykymaailmassa kasvava uhkansa, koska siihen järjestelmään on solutettu tunnettu narkotika, joka estää fysiologisen aivouudistumisen ja synaptisen plastisuuden sekä epätäsmentää kognition niin korkealla tasolla, että henkilöllä ei ole insiktiä molekyylin aiheuttamasta vaarasta. Insiktin muodostuminen vaatii että insiktiä muodostava aivotaso ei ole myrkyttynyt ja siinä huipun huipputasossa on kannabis inkräktare, väärä molekyyli, voisi sanoa, biologinen ase maailman aivon kutistamiseen ja insiktin poistamiseen.
Cannabis estää neuronin uusiutumisen ja kasvun, minkä havaitsee parhaiten tämän järjestelmän tärkeydestä jo embryonaalisessa kehityksessä- (Voisi sanoa että cannabis lienee lähinnä globaali ehkäisymenetelmä, joka perustuu embryon muuttamiseen elinkyvyttömäksi).
Kun kuuntelee niitten johtohenkilöitten puheita, jotka itse käyttävät tuota molekyyliä huomaa täydellisen insiktin puutteen.
Normaalin aivon ajattelun energiapurkaukset toimivat tehokkaalla glutamaattihappo (Glu) ja GABA sekä dopamiinieritysjärjestelmillä, ja näitä kaikkia alkuvoimaerityksia kannabis estää ja säätää niitä mataliksi ja ehkä aaltomaisesti itsestään purkautuviksi, kun varsinainen tahdonalainen alkusysäys alenee ja kuitenkin metabolia sykkii uutta aminohappoa jätetypestä ja aineenvaihdunnasta käsin kertymään johonkin altaaseen.
Cannabisvaikutus on kaotogeeninen ja sisäsyntyisiä hallusinaatioita aiheuttava ja motoriikkaa impulsiiviseksi ja kontrolloimattomaksi tekevää.
Kannabis tulisi kirjata vaarallisiimpiin narkotikoihin, koska insiktin kadottua se avaa tiet kaikille muille narkotikoille.
Asetan tämän rasvahappoblogiin, koska aivomme on lähinnä rasvaaineista, lipideistä koostunut moduli ja tarvitsee puhtaita rasva-aineita ravinnoksi. Kasvisöljyt ovat suotuista ruokaa. Samoin rasvainen kala, joka antaa DHA ja EPA kalarasvoja. Ne tasapainottavat kasvisöljyperäisen ja linolihaposta käsin muodostuvan arakidonihapon tehokasta aiuneenvaihduntaa.
Aivoille on tärkeää että proteiinien ja hiilihydraattien saanti on myös tasapainoista. Aivot ovat suuri rauhanen, joka eritää paljon hormoneja, jotka ovat proteiineja ja aminohappoketjuja. hermonvälittäjäaineet ovat aminohappoja.
Monta kertaa narkotikan käyttäjät tekevät vielä myrkkymolekyylin nauttimisen lisäksi sen virheen, että eivät syö tavallista ruokaa. Alkoholi antaa vain 2 hiilen ketjuja ja on orgaaninen liuotin, mikä lisää kaikkien muitten myrkkyjen vaarallista vaikutusta aivoon.
Endokannabinoidin synteesi ja vapautuminen
Endocannabinoid synthesis & release
- Tavallisessa hermoimpulssin välityksessä presynaptinen hermosolu vapauttaa hermonvälittäjäainetta synaptiseen välitilaan, missä se kiinnittyy omaan reseptoriinsa postsynaptisen hermon puolella.
- Kun sitoutuminen tapahtuu, neuroni depolarisoituu.
- Tämä depolarisoituminen kiihdyttää kalsiumjonin Ca++ sisäänmenoapostsynaptiseen neuroniin.
- Tämä kalsiumin lisä taas aktivoi entsyymin joka on transasylaasi ja se katalysoi ensimmäistä askelta endokannabinoidien biosynteesissä muuttamalla kalvofosfolipidin PE kefaliinin eli fosfatidyylietanolaminin) muotoon NAPE, N-asyyli-fosfoatidyylietanolamini, N-Acyl- kefaliini.
In standard neurotransmission, the pre-synaptic neuron releases neurotransmitter into the synaptic cleft which binds to cognate receptors expressed on the post-synaptic neuron.
Upon binding, the neuron depolarizes.
This depolarization facilitates the influx of calcium into the neuron;
this increase in calcium activates an enzyme called transacylase which catalyzes the first step of endocannabinoid biosynthesis by converting phosphatidylethanolamine, a membrane-resident phospholipid, into N-acyl-phosphatidylethanolamine (NAPE).
- On osoitettu kokeellisesti että monet fosfolipaasit voivat pilkkoa NAPE- muotoa ja silloin tulee anandamidia (AEA)
Experiments have shown that multiple phospholipases cleave NAPE to yield anandamide [8][9].
- Jos NAPE-PLD enstyymi puuttuisi, kuitenkin havaitaan anandamidin (AEA) biosynteesiä, joten muitakin entsyymejä on asialla.
In NAPE-phospholipase D (NAPE-PLD) knockouts, the PLD-mediated cleavage of NAPE is reduced, not abolished, in low calcium concentrations, suggesting multiple, distinct pathways are involved in AEA biosynthesis (Leung et al., 2006).
- Kun endokannabinoidia ( anandamidia, AEA) on päässyt solunulkoiseen tilaan oletetulla endokannabinoidikuljettajalla, nämä lipidiset välittäjäaineet ovat alttiina gliasolutaholta tapahtuvalle inaktivaatiolle.
Once released into the extracellular space by a putative endocannabinoid transporter, messengers are vulnerable to glial inactivation.
- Endokannabinoidit otetaan talteen oletetun kuljettajan kautta ja sitten ne hajoittuvat rasvahappoamidin hydrolaasilla (FAAH), joka pilkkoo anandamidin ja entsyymillä MGLL, joka pilkkoo toisen endokannabinoidin 2-AG ja tuloksena on ensinmainitussa tapauksessa arakidonihappoa(AA) ja etanolaminia (EA) ja jälkimmäisessä tapauksessa arakidonihappoa(AA) ja glyserolia.
Endocannabinoids are uptaken via a putative transporter and degraded by fatty acid amide hydrolase (FAAH) which cleaves anandamide and MGLL, which cleaves 2-AG to arachidonic acid & ethanolamine and arachidonic acid & glycerol, respectively (reviewed in Pazos et al., 2005).
- Nyt on muodostunut saataville toisen entsyymijärjestelmän substraattiaineita: ARAKIDONIHAPPO tunnistetaan leukotrieenejä (LTs) ja prostaglandiineja (PGs) muodostavien entsyymien järjestelmässä muualla kehosoluissa, mutta on ollut kyseenalaista valiutuuko tämä aineenvaihduntareitti myös keskushermostossa ja onko LT ja PG tuotteilla jotain uutta tehtävää löydettävissä aivoalueella
While arachidonic acid is a substrate for leukotriene and prostaglandin synthesis, it is unclear whether this degradative byproduct has novel functions in the CNS (Yamaguchi et al., 2001; Brock, T., 2005).
- On kasvavaa tietoa myös siitä, että FAAH entsyymiä ilmenee postsynaptisessa neuronissa joilloin se osallistuisi anadamidin ja 2-AG molekyylin poissiivoukseen ja inaktivoimiseen takaisinoton kautta.
Emerging data in the field also points to FAAH being expressed in the postsynaptic neuron, suggesting it also contributes to the clearance and inactivation of anandamide and 2-AG by endocannabinoid reuptake.
Tästä asiasta otan eri artikkeleita jotka valaisevat eri näkökohtia.
Endokannabinoidit wikipediassa
The endocannabinoid system refers to a group of neuromodulatory lipids and their receptors that are involved in a variety of physiological processes including appetite, pain-sensation, mood, and memory. It is named for endocannabinoids, the endogenous lipids that bind cannabinoid receptors (the same receptors that mediate the psychoactive effects of cannabis). Broadly speaking, the endocannabinoid system refers to:
- The cannabinoid receptors CB1 and CB2, two G protein-coupled receptors primarily located in the central nervous system and periphery, respectively.
- The endogenous arachidonate-based lipids, anandamide (N-arachidonoylethanolamine, AEA)) and 2-arachidonoylglycerol (2-AG), collectively termed the "endocannabinoids", that are ligands for the cannabinoid receptors.
- Enzymes synthesize and degrade the endocannabinoids anandamide and 2-AG. Unlike neurotransmitters, endogenous cannabinoids are not stored in vesicles after synthesis, but are synthesized on demand (Rodriguez de Fonseca et al., 2004)[citation needed].
The endocannabinoid system has been studied using genetic and pharmacological methods. These studies have revealed a broad role for endocannabinoid signaling in a variety of physiological processes, including neuromodulator release,[1][2][3] motor learning,[4] synaptic plasticity,[5] appetite,[6] and pain sensation.[7]
Contents
Miten jatkuu endokannabinoiditutkimus Ruotsissa?
Sitaatti internetistä vuodelta 2008.
Tibor Harkany forskning handlar om att förstå hur cannabis påverkar hjärnans utveckling. Nu är han utvald att ingå i ett prestigefyllt europeiskt nätverk för unga forskare.
Datum: 2008-04-28
Tibor Harkany forskning handlar om att förstå hur cannabis påverkar hjärnans utveckling. Nu är han utvald att ingå i ett prestigefyllt europeiskt nätverk för unga forskare.
Cannabis verkar via speciella receptorer i nervcellerna i hjärnan. I fjol kunde Tibor Harkany och hans kollegor vid Karolinska institutet rapportera om fyndet av en grupp naturliga ämnen som verkar genom att binda till samma receptor. Upptäckten, som kan förklara hur droger från cannabis- växten påverkar hjärnan, publicerades i tidskriften Science.
Siv Engelmark Cederborg– Nu vill vi visa när och var under nervsystemets utveckling de olika naturliga ämnena, endocannabinoiderna finns, och hur dessa påverkar nervcellerna, säger Tibor Harkany.
Han är speciellt intresserad av hur drogerna påverkar hjärnans utveckling. Tidigare studier har visat att barn vars mor rökt cannabis under fostertiden kan få problem med social interaktion, kognition och inlärning. Men vad som egentligen händer på cellnivå är ännu inte känt. Det ska forskningen på KI visa.
Men Tibor Harkany själv finns numera vid universitetet i Aberdeen, efter sju år i Stockholm.
– Du vill inte lämna KI. Det är en utmanade forskningsmiljö, en av de bästa i Europa. Men i det svenska systemet finns ett gap på fem, sex år efter att man har varit forskarassistent, där det är extremt svårt att hitta en position för en forskare. Har du barn och familj är det inte så lätt, säger han när Tentakel fångar in honom under ett besök i Sverige.
När han blev erbjuden en permanent professorstjänst, med lön på livstid, var beslutet ändå inte svårt. För tio månader sedan gick flyttlasset till Aberdeen.
Aberdeen är historiskt ett center för cannabis- forskning i Europa. Universitets forskare var med bakom upptäckten av den aktiva substansen i drogen för 40 år sedan.
– De vill öka kapaciteten i cell- och molekylärbiologi och har anställt mig, berättar Tibor Harkany.
Nu sitter han vid ett cafébord på KI-området med en kopp kaffe framför sig. Han är här för att se till forskargruppen vid avdelningen för medicinsk biokemi och biofysik, som han fortfarande leder. I överenskommelsen med universitetet i Aberdeen ingår att han får fortsätta jobba vid KI när han behöver.
Här använder han institutets nya elektrofysiologi-labb för att studera hur nervceller integrerar och kommunicerar med varandra i musembryon och human fostervävnad. Han tittar också på hur kommunikationen påverkas av cannabis.
Tibor Harkany plockar bland kaffemuggarna på cafébordet för att förklara. Locken på muggarna får bli nervceller. Träskeden förbinder dem, liksom celler som utvecklas normalt binds samman i ett visst mönster.
Hans hypotes är att cannabis leder nervcellerna fel så att de inte kopplas ihop som de ska hos det växande fostret. Därmed påverkas utvecklingen av fostrets hjärna.
– Vi tror att cannabis avbryter tillväxten av utskotten som binder samman nervcellerna, säger han.
Nu har han som en av 18 unga forskare bland 200 sökande valts ut till det europeiska programmet EMBO Young Investigator. Det ger bland annat ett nätverk med andra forskare. Från Vetenskapsrådet kommer 192 000 kronor per år under tre år, som kan användas till resor, utrustning, kemikalier.
– Utnämningen öppnar nya dörrar och ger mig unika möjligheter till samarbeten och kontakter. Det är en bekräftelse på mina forskningsansträngningar och hårt arbete under 15 år.
Namn: Tibor Harkany. 36 år.
född i Ungern och är forskarutbildad i Ungern och Groningen i Nederländerna.
Datum: 2008-04-28
Prenumerera på:
Kommentarer (Atom)
{kind=link}