http://www.avoin.helsinki.fi/oppimateriaalit/ravitsemustieteen_perusteet/04_rasvat_rakenne.shtml
Tässä ravitsemustieteen perusteessa huomaa rasvahappojen joukossa kasvisperäisen linolihapon.
Siitä muodostuu kehossa arakidonihappoa, jota joka solu lataa fosfolipideihinsä ja kalvoihinsa.
Siitä on sitten paikallisesti purettavissa takaisin esiin arakidonihappoa, jota taas entsyymisetti modifioi työkaluksi paikallisiin tehtäviin, immunologisiin, reaktiivisiin, tulehduksellisiin, signaloiviin ym tarpeista riippuen.
Koska arakidonihapossa ( eikosatetraeenihapossa) on 20 hiiltä (eicosa), niin sen johdannaisia sanotaan eikosanoideiksi.
Tetraeeni tarkoittaa että siinä on 4 kpl kaksoissidoksia rakenteessa.
Eri entsyymiseteistä sitten lähtee käyntiin eikosanoidikaskadit. Ne ovat myös kudosspesifisiä ( verisuonten puolella, keuhkoissa, ihossa, eri solukalvoissa, valkosoluissa, aivoissa jne on omat tyyppireaktionsa ja omat molekyylinsä).
PG= prostaglandiinit
LT= leukotrieenit
LX= lipoxygenaasit
HPETE=hydroperoxieikosatetraeenihappo
http://upload.wikimedia.org/wikipedia/commons/thumb/4/40/Eicosanoid_synthesis.svg/400px-Eicosanoid_synthesis.svg.png
Mitä kaikkea vapaasta arakidonihaposta voi tulla mm?
http://groundupstrength.wdfiles.com/local--files/health:omega-3-fatty-acids-and-inflammation/eicosanoid%20biosynthesis.JPG
Tämä on tosi aggressiivisten reaktiivisten molekyylien puu.
Ne kovasti mainostetut kalarasvat muodostavat näitten sukuisia molekyylejä ,jotka ovat vähemmän aggressiivisia ja täten vaimentavat ja miedontavat reaktiivisen arakidonihapon eikosanoidikaskadia.
Kts. Eikosanoidi keuhkofibroosissa
http://www.clinsci.org/cs/108/cs1080479.htm#CitedBy
lördag 11 december 2010
{kind=link}
{kind=link}
Aspiriini moduloi rasvahappoaineenvaihduntaa
http://scienceblog.cancerresearchuk.org/2010/12/07/aspirin-cuts-risk-of-dying-from-several-types-of-cancer/
JOS niin on että aspiriini on edullinen monia syövän kehittymisiä vastaan, niin mikä aiheuttaa syövän? Aspiriini jarruttaa arakidonihappoaineenvaihdunnan eikosanoidikaskadia, mutta ei sen kaikkia teitä.
http://jcp.bmj.com/content/57/1/6/F1.large.jpg
http://img.medscape.com/fullsize/migrated/405/963/ch1185.02.lieb.fig1.gif
Ei voi sanoa edes niin, että arakidonihapon vähempi saatavuus olisi edullisempaa, sillä sen normaali saatavuus pitää eikosanoidijärjestelmän kaikki säätelyt tasapainossa.
Arakidonihapon puute taas on hyvin paha tilanne, missä ihmisen solut eivät korjaannu. Arakidonihappoa asettuu kaikkiin solukalvoihin koko kehossa. Se on linolihappoperäistä.
Eikosandoi ja solukorjautumisjärjestelmän suistuminen tulehdukselliselle tielle ja tulehduksen jatkuminen nukleaariseen tasoon, solun muuttumiseen syöpäsoluksi, täytyy olla jokin rasvahappo ym aineenvaihdunnan poikkeusmolekyyli syynä eikä normaalin eikosanoidikaskadin kuvan kauniit molekyylit. MUTTA toisaalta, eikosanoidijärjestelmän työmolekyylien valmistuksessa tulee aika runsasti niitä sivurakenteita ja niistä käsin voi luonnollisesti löytyä karsinogeenejä, jos ne eivät häviä omissa silppureissaan.
JOS niin on että aspiriini on edullinen monia syövän kehittymisiä vastaan, niin mikä aiheuttaa syövän? Aspiriini jarruttaa arakidonihappoaineenvaihdunnan eikosanoidikaskadia, mutta ei sen kaikkia teitä.
http://jcp.bmj.com/content/57/1/6/F1.large.jpg
{kind=link}
http://img.medscape.com/fullsize/migrated/405/963/ch1185.02.lieb.fig1.gif
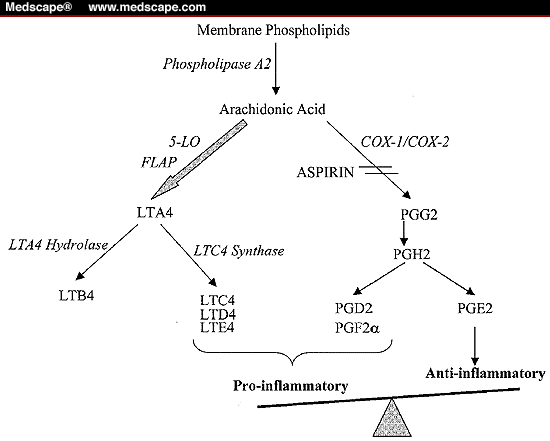{kind=link}
Ei voi sanoa edes niin, että arakidonihapon vähempi saatavuus olisi edullisempaa, sillä sen normaali saatavuus pitää eikosanoidijärjestelmän kaikki säätelyt tasapainossa.
Arakidonihapon puute taas on hyvin paha tilanne, missä ihmisen solut eivät korjaannu. Arakidonihappoa asettuu kaikkiin solukalvoihin koko kehossa. Se on linolihappoperäistä.
Eikosandoi ja solukorjautumisjärjestelmän suistuminen tulehdukselliselle tielle ja tulehduksen jatkuminen nukleaariseen tasoon, solun muuttumiseen syöpäsoluksi, täytyy olla jokin rasvahappo ym aineenvaihdunnan poikkeusmolekyyli syynä eikä normaalin eikosanoidikaskadin kuvan kauniit molekyylit. MUTTA toisaalta, eikosanoidijärjestelmän työmolekyylien valmistuksessa tulee aika runsasti niitä sivurakenteita ja niistä käsin voi luonnollisesti löytyä karsinogeenejä, jos ne eivät häviä omissa silppureissaan.
torsdag 4 november 2010
Endokannabinoidisysteemi endokriinisessä säätelyssä ja energiatasapainossa
Endocrine Reviews 27 (1): 73-100 The Emerging Role of the Endocannabinoid System in Endocrine Regulation and Energy Balance
Endocrinology Unit, Department of Internal Medicine and Gastroenterology et al.
During the last few years, the endocannabinoid system has emerged as a highly relevant topic in the scientific community. Endokannabinoidien nimiin on jo kirjattu monia erilaisia säätelytoimintoja ja niitten osuutta useissa patofysiologisissa tiloissa tutkitaan intensiivisesti. Many different regulatory actions have been attributed to endocannabinoids, and their involvement in several pathophysiological conditions is under intense scrutiny. Kannabinoidireseptorit CB1 ja CB2 löydettiin alunperin Cannabis sativa-nimisen kasvin psykotrooppisen komponentin ( cannabinoidien) reseptoreina. Nämä reseptorit osallistuvat monien keskushermoston ja perifeerisen hermoston fysiologisten funktioitten säätelyyn. Cannabinoid receptors, named CB1 receptor and CB2 receptor, first discovered as the molecular targets of the psychotropic component of the plant Cannabis sativa, participate in the physiological modulation of many central and peripheral functions. CB2 reseptoria esiintyy lähinnä immuunisoluissa, kun taas CB1 reseptori on runsain G-proteiiniin kytkeytynyt reseptori, mitä aivoissa ilmenee. CB2 receptor is mainly expressed in immune cells, whereas CB1 receptor is the most abundant G protein-coupled receptor expressed in the brain. CB1 reseptoria ilmenee hypotalamuksessa ja hypofyysissä. Tiedetään, että sen aktivaatio moduloi kaikkia endokriinisia hypotalamo-perifeerisiä endokriinisia akseleita. CB1 receptor is expressed in the hypothalamus and the pituitary gland, and its activation is known to modulate all the endocrine hypothalamic-peripheral endocrine axes. On lisääntyvää näyttöä, että tämä EC-järjestelmä omaa osuuden stressivasteessa vaikuttamalla hypotalamus-hypofyysi-lisämunuais-akseliin ja reproduktion kontrollissa modifioimalla gonadotropiinien vapautumista, fertiliteettiä ja seksuaalista käyttäytymistä. An increasing amount of data highlights the role of the system in the stress response by influencing the hypothalamic-pituitary-adrenal axis and in the control of reproduction by modifying gonadotropin release, fertility, and sexual behavior.Endokannabinoidijärjestelmän (EC) kyky kontrolloida ruokahalua, ravinnonottoa ja energiatasapainoa on saanut viime aikoina myös suurta huomiota erityisesti näiden funktioiden taustatekijöiden suhteen. The ability of the endocannabinoid system to control appetite, food intake, and energy balance has recently received great attention, particularly in the light of the different modes of action underlying these functions. Endokannabinoidijärjestelmä moduloi ravinnon palkitsevaisuutta vaikuttamalla erityisiin mesolimbisiin alueisiin aivoissa. The endocannabinoid system modulates rewarding properties of food by acting at specific mesolimbic areas in the brain. In the hypothalamus, CB1 receptor and endocannabinoids are integrated components of the networks controlling appetite and food intake. Mielenkiintoista on , että endokannabinoidien on viime aikoina osoitettu kontrolloivan aineenvaihdunnallisia funktioita perifeerisissä kudoksissa kuten adiposyyteissä, maksasoluissa, mahasuolikanavassa ja mahdollisesti tahdonalaisessa lihaksessa. Interestingly, the endocannabinoid system was recently shown to control metabolic functions by acting on peripheral tissues, such as adipocytes, hepatocytes, the gastrointestinal tract, and, possibly, skeletal muscle. Järjestelmän relevanssia vahvistaa edelleen huomio siitä, että ECV-järjestelmään interaktiossa olevien lääkkeiden on havaittu olevan lupaavia kandidaatteja erilaisten sairauksien hoitoon, joihin luetaan myös liikalihavuus. The relevance of the system is further strenghtened by the notion that drugs interfering with the activity of the endocannabinoid system are considered as promising candidates for the treatment of various diseases, including obesity. |
Anadamidi ja dopaminerginen neuroni
- Anadamidi ja DA neuroni
Tässä on kuva neuronista joka on dopaminerginen. Tämän kuvan laitoin pohdintoja varten. Tämä on hahmoteltu vuonna 1999.
- Anadamidihermotuksen kliinisestä merkityksestä. Pohdintaa varten (ADHD alue) Tässä tutkimuksessa oli nostettu EPA osuutta PUFA hapoissa ja vaste oli neurologisesti edullinen ADHD:ssa. Supplementit, joissa on korkea annos EPA/DHA konsentraatteja voivat parantaa ADHD lasten käytösongelmia. Tämä artikkeli ei suoraan mainitse anadamidin osuutta, mutta sillä on epäsuorasti merkittävä osuus, kunhan sen saa selville.
CONCLUSION: The findings of this small pilot study suggest supplementation with high-dose EPA/DHA concentrates may improve behavior in children with ADHD.
Effects of an open-label pilot study with high-dose EPA/DHA concentrates on plasma phospholipids and behavior in children with attention deficit hyperactivity disorder. Sorgi PJ, Hallowell EM, Hutchins HL, Sears B. Nutr J. 2007 Jul 13;6:16.PMID: 17629918 [PubMed - indexed for MEDLINE]Free PMC ArticleFree text
EPA ( C20:5 n6) and anandamide
J Nutr Biochem. 2010 Oct 14. Eicosapentaenoic acid decreases expression of anandamide synthesis enzyme and cannabinoid receptor 2 in osteoblast-like cells. Hutchins HL, Li Y, Hannon K, Watkins BA. Lipid Chemistry and Molecular Biology Laboratory, Purdue University, West Lafayette, IN 47907, USA.
Abstract (Suomennosta)
ANANDAMIDI (AEA) on endogeeninen agonisti CB2 reseptorille, jota ilmenee osteoblasteissa.
Anandamide (AEA) is an endogenous agonist for the cannabinoid receptor 2 (CB2) which is expressed in osteoblasts.
ARAKIDONIHAPPO (AA) on anandamidin prekursoriaine; ravintoperäisen n3- linjaisen PUFA rasvahapon tiedetään vähentävän arakidonihapon pitoisuutta kudoksissa ja soluissa.
Arachidonic acid (AA) is the precursor for AEA and dietary n-3 polyunsaturated fatty acids (PUFA) are known to reduce the concentrations of AA in tissues and cells.
- Hypoteesi
Tästä tutkijaryhmä teki hypoteesinsa siitä, että n3 PUFA hapot, eikosapentaeenihappo (EPA) ja dokosaheksaeenihappo (DHA), jotka vähentävät solujen arakidonihappopitoisuuksia, voisivat alentaa anandamidin pitoisuutta osteoblasteissa muuntamalla endokannabinoidisysteemin entsyymien ilmenemistä.
Therefore, we hypothesized that n-3 PUFA, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which reduce AA in cells, could lower AEA in osteoblasts by altering enzyme expression of the endocannabinoid (EC) system.
- (Tekniikasta selitystä:
MC3T3-E1 osteoblast-like cells were grown for 6, 10, 15, 20, 25 or 30 days in osteogenic medium. Osteoblasts were treated with 10 μM of AA, EPA, DHA, oleic acid (OA) or EPA+DHA (5 μM each) for 72 h prior to their collection for measurement of mRNA and alkaline phosphatase (ALP) activity.)
- PUFA n6 ja n3 linjoista
Osteoblastit, joita käsiteltiin arakidonihapolla sisälsivät enemmän arakidonihappoa ja n6- linjan PUFA happoja kuin ne joita käsiteltiin EPA ja DHA-hapoilla, joilla oli matalampi n6 linjan kuin n3- linjan happojen pitoisuus
Compared to vehicle control, osteoblasts treated with AA had higher levels of AA and n-6 PUFA while those treated with EPA and DHA had lower n-6 but higher n-3 PUFA.
- Tuloksista
Riippumatta rasvahappokäsittelystä osteoblastit kypsyivät normaalisti, mikä ilmeni ALP-entsyymistä.
Independent of the fatty acid treatments, osteoblasts matured normally as evidenced by ALP activity.
- NAPE-PLD, FAAH, CB2
NAPE-selektiivisen PLD-entsyymin, rasvahappoamidihydrolaasin (FAAH) ja CB2mRNA:n ilmenemät olivat suuremmat 20 päivän kuin 10 päivänä tuloksissa.
N-acyl phosphatidylethanolamine-selective phospholipase D (NAPE-PLD), fatty acid amide hydrolase (FAAH) and CB2 mRNA expression were higher at 20 days compared to 10 days.
NAPE-PLD ja CB2mRNA ilmenemät olivat matalammat niissä osteoblasteissa, joita oli käsitelty EPA rasvahapolla verrattuna kaikkiin muihin ryhmiin.
NAPE-PLD and CB2 mRNA was lower in osteoblasts treated with EPA compared to all other groups.
Täten havaitaan, että NAPE-PLD mRNA, FAAH mRNA ja CB2mRNA lisääntyivät osteoblastin kypsymisen aikana ja että EPA rasvahappo vähensi NAPE-PLD entsyymin ja CB2 reseptorin mRNA:ta
Thus, mRNA expression for NAPE-PLD, FAAH, and CB2 increased during osteoblast maturation and EPA reduced mRNA for NAPE-PLD and CB2 receptor.
- Yhteenveto
EPA alensi endokannabinoidisysteemin(EC) proteiinien mRNA pitoisuuksia ja anandamidiaineenvaíhdunnan synteesivaiheen ja hajoamisvaiheen aineenvaihdunnan entsyymien mRNA-pitoisuuksia osteoblasteissa.
In conclusion, EPA lowered mRNA levels for proteins of the EC system and mRNA for AEA synthesis/degradation is reported in osteoblasts.
LinkOut - more resources
söndag 31 oktober 2010
FOSFOLIPIDI
Solujen kalvot ovat fosfolipidien kaksoiskerroksia.
Fosfolipidi (PL) sijoittaa glyserolirakneteiseen hyllymäiseen sijoitukseen tärkeitä rasvahappoja, joita tarvitaessa ( on demand) hyödynnetään eri puolilla kehoa. Yksi näistä kaikkiin soluihin sijoitetusta rasvahaposta on arakidonihappo (AA), jota on saatavilla tarpeen mukaan ja se osallistuu kehon sorvauksiin ja plastisuuden ylläpitämisiin.
Types of phospholipid, fosfolipidityypit
Diacylglyceride structures, diglyseridirakenteet
Phosphosphingolipids, Fosfosfingolipidit
Phospholipid derivatives, fosfolipidijohdannaiset
On geneettisesti ohjautunutta mihin asemaan minkäkinlainen rasvahappo mieluiten asettuu , essentielleillä rasvahapoilla (EFA) on erityinen prioriteettinsa. Jos niitä ei ole valiutuu non-essentiellejä näille paikoille.
Essentiellit rasvahapot linolihappo ja linoleenihappo pystyvät kehoentsyymien kautta muodostamaan sellaisia rasvahappoja, joihin kehon korkea-asteisesti spesifiset entsyymijärjestelmät voivat pureutua ja käyttää näitä molekyyleja tarkoin säädeltyinä työaseina kehon hoidossa, kuten esim avaruusasemaa hoidettaessa, kun korjataan jotain modulia ulkoavaruuden puolella ja käytetään jotain työasetta kädessä.
Arakidonihappojärjestelmä on tällainen huipputäsmällinen työase kehon kunnossapidossa. Se otetaan esiin näistä rakenteellisista säilöpaikosita rakenteellisista lipideistä ja muokataan soveltuvaksi työaseeksi.
Tällainen työase otetaan vain tarvittaessa esiin, "on demand" ja hoidetaan sitten inaktiiviseen muotoon takaisin työn jälkeen tavalla tai toisella.
Koska linolihapoista muodostuva ARAKIDONIHAPPO on aika aggressiivinen aine sen johdannaisten muodostuttua, linoleenihaposta käsin on sitä järejstelmäkin olemassa paralleelisti. sofistisesti vaimentavaLinoleenihappolinjasta tunnetaan EPA ja DHA joita voi saada vlamiissakin muodossa kalarasvoista käsin.
Fosfolipidi (PL) sijoittaa glyserolirakneteiseen hyllymäiseen sijoitukseen tärkeitä rasvahappoja, joita tarvitaessa ( on demand) hyödynnetään eri puolilla kehoa. Yksi näistä kaikkiin soluihin sijoitetusta rasvahaposta on arakidonihappo (AA), jota on saatavilla tarpeen mukaan ja se osallistuu kehon sorvauksiin ja plastisuuden ylläpitämisiin.
Types of phospholipid, fosfolipidityypit
Diacylglyceride structures, diglyseridirakenteet
- See: Glycerophospholipid
- Phosphatidic acid (phosphatidate) (PA)
- Phosphatidylethanolamine (cephalin) (PE)
- Phosphatidylcholine (lecithin) (PC)
- Phosphatidylserine (PS)
- Phosphoinositides:
- Phosphatidylinositol (PI)
- Phosphatidylinositol phosphate (PIP)
- Phosphatidylinositol bisphosphate (PIP2) and
- Phosphatidylinositol triphosphate (PIP3).
Phosphosphingolipids, Fosfosfingolipidit
- Ceramide phosphorylcholine (Sphingomyelin) (SPH)
- Ceramide phosphorylethanolamine (Cer-PE)
- Ceramide phosphorylglycerol
Phospholipid derivatives, fosfolipidijohdannaiset
- See table below for an extensive list.
- Natural phospholipid derivates:
- egg PC, egg PG, soy PC, hydrogenated soy PC, sphingomyelin as natural phospholipids.
- Synthetic phospholipid derivates:
- Phosphatidic acid (DMPA, DPPA, DSPA)
- Phosphatidylcholine (DDPC, DLPC, DMPC, DPPC, DSPC, DOPC, POPC, DEPC)
- Phosphatidylglycerol (DMPG, DPPG, DSPG, POPG)
- Phosphatidylethanolamine (DMPE, DPPE, DSPE DOPE)
- Phosphatidylserine (DOPS)
- PEG phospholipid (mPEG-phospholipid, polyglycerin-phospholipid, funcitionalized-phospholipid, terminal activated-phospholipid)
Abbreviations used and chemical information of glycerophospholipids
Abbreviation | CAS | Name | Type |
|---|---|---|---|
| DDPC | 3436-44-0 | 1,2-Didecanoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DEPA-NA | 80724-31-8 | 1,2-Dierucoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DEPC | 56649-39-9 | 1,2-Dierucoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DEPE | 988-07-2 | 1,2-Dierucoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DEPG-NA | 1,2-Dierucoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol | |
| DLOPC | 998-06-1 | 1,2-Dilinoleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DLPA-NA | 1,2-Dilauroyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid | |
| DLPC | 18194-25-7 | 1,2-Dilauroyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DLPE | 1,2-Dilauroyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine | |
| DLPG-NA | 1,2-Dilauroyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol | |
| DLPG-NH4 | 1,2-Dilauroyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol | |
| DLPS-NA | 1,2-Dilauroyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DMPA-NA | 80724-3 | 1,2-Dimyristoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DMPC | 18194-24-6 | 1,2-Dimyristoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DMPE | 988-07-2 | 1,2-Dimyristoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DMPG-NA | 67232-80-8 | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DMPG-NH4 | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol | |
| DMPG-NH4/NA | 1,2-Dimyristoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium/Ammonium Salt) | Phosphatidylglycerol | |
| DMPS-NA | 1,2-Dimyristoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DOPA-NA | 1,2-Dioleoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid | |
| DOPC | 4235-95-4 | 1,2-Dioleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DOPE | 4004-5-1- | 1,2-Dioleoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DOPG-NA | 62700-69-0 | 1,2-Dioleoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DOPS-NA | 70614-14-1 | 1,2-Dioleoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine |
| DPPA-NA | 71065-87-7 | 1,2-Dipalmitoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DPPC | 63-89-8 | 1,2-Dipalmitoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DPPE | 923-61-5 | 1,2-Dipalmitoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DPPG-NA | 67232-81-9 | 1,2-Dipalmitoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DPPG-NH4 | 73548-70-6 | 1,2-Dipalmitoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol |
| DPPS-NA | 1,2-Dipalmitoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| DSPA-NA | 108321-18-2 | 1,2-Distearoyl-sn-glycero-3-phosphate (Sodium Salt) | Phosphatidic acid |
| DSPC | 816-94-4 | 1,2-Distearoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| DSPE | 1069-79-0 | 1,2-Distearoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine |
| DSPG-NA | 67232-82-0 | 1,2-Distearoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Sodium Salt) | Phosphatidylglycerol |
| DSPG-NH4 | 108347-80-4 | 1,2-Distearoyl-sn-glycero-3[Phospho-rac-(1-glycerol...) (Ammonium Salt) | Phosphatidylglycerol |
| DSPS-NA | 1,2-Distearoyl-sn-glycero-3-phosphoserine (Sodium Salt) | Phosphatidylserine | |
| Egg Sphingomyelin empty Liposome | |||
| EPC | Egg-PC | Phosphatidylcholine | |
| HEPC | Hydrogenated Egg PC | Phosphatidylcholine | |
| HSPC | High purity Hydrogenated Soy PC | Phosphatidylcholine | |
| HSPC | Hydrogenated Soy PC | Phosphatidylcholine | |
| LYSOPC MYRISTIC | 18194-24-6 | 1-Myristoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| LYSOPC PALMITIC | 17364-16-8 | 1-Palmitoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| LYSOPC STEARIC | 19420-57-6 | 1-Stearoyl-sn-glycero-3-phosphocholine | Lysophosphatidylcholine |
| Milk Sphingomyelin MPPC | 1-Myristoyl-2-palmitoyl-sn-glycero 3-phosphocholine | Phosphatidylcholine | |
| MSPC | 1-Myristoyl-2-stearoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| PMPC | 1-Palmitoyl-2-myristoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| POPC | 26853-31-6 | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
| POPE | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine | Phosphatidylethanolamine | |
| POPG-NA | 81490-05-3 | 1-Palmitoyl-2-oleoyl-sn-glycero-3[Phospho-rac-(1-glycerol)...] (Sodium Salt) | Phosphatidylglycerol |
| PSPC | 1-Palmitoyl-2-stearoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| SMPC | 1-Stearoyl-2-myristoyl-sn-glycero-3–phosphocholine | Phosphatidylcholine | |
| SOPC | 1-Stearoyl-2-oleoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine | |
| SPPC | 1-Stearoyl-2-palmitoyl-sn-glycero-3-phosphocholine | Phosphatidylcholine |
- PC tarkoittaa fosfatidyylikoliineja, lesitiinejä
- PS tarkoittaa fosfatidyyliseriinejä
- PE tarkoittaa fosfatidyylietanolamineja, kefaliineja
- PI tarkoittaa lipositoleja, fosfoinositolilajeja
- LPA tarkoittaa lysolesitiiniä, lysofsofatidyylikoliineja
- PA tarkoittaa fosfatidihappoa
- Ptd-glyserolikin on listoissa
On geneettisesti ohjautunutta mihin asemaan minkäkinlainen rasvahappo mieluiten asettuu , essentielleillä rasvahapoilla (EFA) on erityinen prioriteettinsa. Jos niitä ei ole valiutuu non-essentiellejä näille paikoille.
Essentiellit rasvahapot linolihappo ja linoleenihappo pystyvät kehoentsyymien kautta muodostamaan sellaisia rasvahappoja, joihin kehon korkea-asteisesti spesifiset entsyymijärjestelmät voivat pureutua ja käyttää näitä molekyyleja tarkoin säädeltyinä työaseina kehon hoidossa, kuten esim avaruusasemaa hoidettaessa, kun korjataan jotain modulia ulkoavaruuden puolella ja käytetään jotain työasetta kädessä.
Arakidonihappojärjestelmä on tällainen huipputäsmällinen työase kehon kunnossapidossa. Se otetaan esiin näistä rakenteellisista säilöpaikosita rakenteellisista lipideistä ja muokataan soveltuvaksi työaseeksi.
Tällainen työase otetaan vain tarvittaessa esiin, "on demand" ja hoidetaan sitten inaktiiviseen muotoon takaisin työn jälkeen tavalla tai toisella.
Koska linolihapoista muodostuva ARAKIDONIHAPPO on aika aggressiivinen aine sen johdannaisten muodostuttua, linoleenihaposta käsin on sitä järejstelmäkin olemassa paralleelisti. sofistisesti vaimentavaLinoleenihappolinjasta tunnetaan EPA ja DHA joita voi saada vlamiissakin muodossa kalarasvoista käsin.
Endokannabinoiditiedemiehet
Sign-in free and Explore the Exciting World of BiomedExperts:
- Over 1,800,000 Profiles
- More than 3,500 Organizations worldwide
- State of the Art Network Visualizations
- Manage your own Profile
- Locate Experts in your Country/Region
- Locate Experts in your 1. and 2. Level Network
- Connect to Experts Worldwide
NetworkView

Experts in Endocannabinoids
Endogenous compounds that bind to CANNABINOID RECEPTORS. They are structurally different from the CANNABINOIDS which are primarily derived from plants.
Prenumerera på:
Kommentarer (Atom)
{kind=link}